Cinclidae, Turdidae, and Muscicapidae
The dippers (Cinclidae) seem to come first, making Turdidae and Muscicapidae sisters (see Barker et al., 2004; Beresford et al., 2005; Treplin et al., 2008; but for a contrary view see Ericson and Johansson, 2003; Voelker and Spellman, 2004).
The remainder of the Muscicapoidea is the Turdidae-Muscicapidae clade, containing almost 500 species. This whole group and its relation to the Sylviioidea has long been contentitous. Ornithologists had great difficulty finding clear-cut ways to distinguish the various groups. This remained true until the DNA era. E.g., the 6th edition AOU checklist includes Sylviinae, Muscicapinae, Monarchinae, Turdinae, and Timaliinae as subfamilies of Muscicapidae! It was also typical to put old world robins, chats and wheatears in the Turdidae (or Turdinae), and various flycatchers in Muscicapidae (Muscicapinae). Sibley and Ahlquist (1990) and Sibley and Monroe (1990) began the process of untangling them by separating the true thrushes in Turdinae and reconsituting Muscicapinae as containing a flycatcher group (Muscicapini) and a robin-chat-wheatear group (Saxicolini). They didn't get it entirely right, but the current arrangement still has these general features. However, the membership of each group has been altered. Note that the Sibley and Monroe subfamilies Muscicapinae and Turdinae are treated as full families, and their tribes have been promoted to subfamilies.
Although the general arrangement of Sibley, Ahlquist, and Monroe has continued to be supported (e.g., Cibois and Cracraft, 2004; Voelker and Spellman, 2004; Zuccon and Ericson, 2010c; Sangster et al. 2010), the details have changed. Indeed, there's been substantial internal restructuring in both the Turdidae and Muscicapidae, with new genera appearing and old genera disappearing under the onslaught of the DNA-based studies.
Cibois and Cracraft already found a Cercotrichas Scrub-Robin and Copsychus Shama grouped in Muscicapinae, while a Ficedula flycatcher ended up with the Saxicolinae. Voelker and Spellman found much the same, and additionally provided a list of taxa erroneously placed in Turdidae, Muscicapinae, and Saxicolinae. Further work has given us a more detailed framework, but within that framework there are a number of surprises and apparently conflicting results.
Dippers
Cinclidae: Dippers Sundevall, 1836
1 genus, 5 species HBW-10
- White-throated Dipper, Cinclus cinclus
- Brown Dipper, Cinclus pallasii
- American Dipper, Cinclus mexicanus
- White-capped Dipper, Cinclus leucocephalus
- Rufous-throated Dipper, Cinclus schulzii
Turdidae: Thrushes Rafinesque, 1815
19 genera, 192 species HBW-10
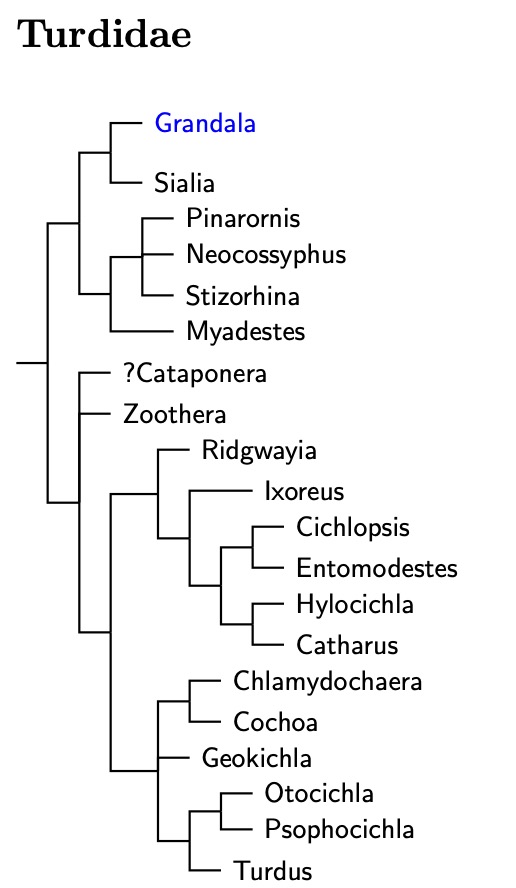 The overall organization of the Turdidae is based on Pasquet et al.
(1999), Klicka et al. (2005), Voelker and Klicka (2008), and
Fjeldså et al. (2020). The
arrangement of Myadestes follows Miller et al. (2007).
Voelker and Klicka (2008) covers the true Zoothera.
Grandala is included in the Turdidae based on Jønsson and
Fjeldså (2006a), Price et al. (2014), and Fjeldså et al. (2020).
The latter also placed the Boulder Chat, Pinarornis plumosus in
a clade with the ant-thrushes (Neocossyphus) and
flycatcher- or rufous-thrushes (Stizorhina).
The overall organization of the Turdidae is based on Pasquet et al.
(1999), Klicka et al. (2005), Voelker and Klicka (2008), and
Fjeldså et al. (2020). The
arrangement of Myadestes follows Miller et al. (2007).
Voelker and Klicka (2008) covers the true Zoothera.
Grandala is included in the Turdidae based on Jønsson and
Fjeldså (2006a), Price et al. (2014), and Fjeldså et al. (2020).
The latter also placed the Boulder Chat, Pinarornis plumosus in
a clade with the ant-thrushes (Neocossyphus) and
flycatcher- or rufous-thrushes (Stizorhina).
The relationships within Turdus itself are still not entirely settled. The problem is that genus has rapidly radiated, with at least 85 species forming in about 7 million years (not including potential splits in the Island Thrush). In such cases, species have often not fully separated before new species are formed, leaving a messy genetic history to sort out. As of 2020, I'm relying on the framework of Batista et al. (2020). However, they didn't include all species. Indeed, their coverage of Old World species is particularly spotty. I have supplemented it using Avendaño et al. (2017) for the Black-billed Thrush complex, Bowie et al. (2005) for Olive Thrushes, Melo et al. (2010) for Gulf of Guinea thrushes. I've also used Nagy et al. (2019), Nylander et al. (2008), and Voelker et al. (2007) as seemed appropriate to fill in. In a few cases I had to guess.
Klicka et al. identified several major clades in Turdidae, but I have not translated them into subfamilies and tribes, mainly because I'm not sure how they fit together. The basal group includes the bluebirds and solitaires. After that come the Australasian and Oriental Zoothera. A big chunk of what had been Zoothera is not closely related to the other part. The Afroasiatic Zoothera are now Geokichla (for details, see Voelker and Outlaw, 2008). The true Zoothera form another clade, as does Catharus and allies. The Sulawesi endemic Geomalia has been move into Zoothera (Olsson and Alström, 2013). There is also a small clade consisting of Chlamydochaera and Cochoa which are allied to Geokichla and Turdus.
There is a split in Zoothera. Based on Alström et al. (2016), the Plain-backed Thrush, Zoothera mollissima, has been split into:
- Alpine Thrush, Zoothera mollissima
- Himalayan Thrush, Zoothera salimalii
- Sichuan Thrush, Zoothera griseiceps
There is probably another species here, “Yunnan Thrush”, but futher work is necessary to confirm this.
Voelker et al. (2013) have analyzed the Catharus clade (Ridgwayia–Catharus) using 2 mitochondrial and 8 nuclear genes. The current arrangement reflects their results and those of Everson et al. (2019). Voelker et al. estimated that the entire clade is about 8 million years old, and that the genus Catharus is about 7 million years. Further, the Veery, Bicknell's, Gray-cheeked clade seems to be about 800,000 years old, with the Bicknell's/Gray-cheeked split about half that age.
As part of the Turdus reorganization, Cichlherminia, Platycichla, and Nesocichla have been merged into Turdus. The genera Myophonus, Brachypteryx, Heinrichia and Alethe have moved to Muscicapidae. Grandala has moved the other way, from Muscicapidae to Turdidae (Jønsson and Fjeldså, 2006a).
I had previously included the Groundscraper Thrush, Psophocichla litsitsirupa, in Turdus, but based on Nagy et al. (2019) and Nylander et al. (2008), I have separated it and the Chinese Thrush from Turdus. I've followed HBW Checklist in putting the Chinese Thrush in Otocichla. It seems most likely to be a distant sister to Psophocichla (see Nagy et al.). I've also followed the HBW Checklist in splitting the Ethiopian Thrush, Psophocichla simensis, from the Groundscraper Thrush, Psophocichla litsitsirupa.
The new arrangement of Turdus has the Mistle and Song Thrushes as basal branches. The remainder is divided into three clades: an African clade, a Eurasian clade, and finally, an American clade.
Based on Nylander et al. (2008), the Chinese Blackbird, Turdus mandarinus, including the subspecies sowerbyi, but not intermedius, has been split from the Eurasian Blackbird, Turdus merula, and the Taiwan Thrush, Turdus niveiceps, has been split from the Island Thrush, Turdus poliocephalus.I've also done some rearrangement of the Olive Thrush complex. Nylander et al. placed the Chinese Blackbird next part of the Olive Thrush complex, but I have some skepticism about that, and put it next to the Eurasian Blackbird. The newly discovered Varzea Thrush, Turdus sanchezorum, has also been added to the list (O'Neill et al., 2011).
Based on Cequeira et al. (2016) and Avendaño et al. (2017), the Pantepui Thrush, Turdus murinus, and the Campina Thrush, Turdus arthuri, have been split from the Black-billed Thrush, Turdus ignobilis.
It is clear from several sources (e.g., Voelker et al., 2007; Nylander et al., 2008; Cerqueira et al., 2016; Avendaño et al, 2017) that Turdus subalaris and Turdus nigriceps are not close relatives, so Slaty Thrush, Turdus nigriceps, is split into Eastern Slaty Thrush, Turdus subalaris, and Andean Slaty Thrush, Turdus nigriceps. I'm not fond of the names, but that is what IOC and the HBW Checklist (del Hoyo and N.J. Collar, 2016) use.
Finally, I have also split Dagua Thrush, Turdus daguae, from the White-throated Thrush, Turdus assimilis (Ridgely and Greenfield, 2001; Nagy et al., 2019; IOC).
The whole Great Thrush complex (serranus, olivater, nigriceps, fuscater, chiguanco) needs further sorting out.
If you examine the Turdus tree, you will find several taxa tagged with a question mark or two. The only genetic information on the Yemen Thrush, Turdus menachensis, is from Nagy et al. (2019), which grouped it with the West Antillean thrushes. The Bare-eyed Thrush, Turdus tephronotus, is near the Mountain Thrush according to Nagy et al., while Nylander et al. (2008) have it grouped with the African thrushes, with the Chinese Blackbird, Turdus mandarinus, basal to both. I have put the first two with the African thrushes, and located the Chinese Blackbird next to the Eurasian Blackbird, although I have no reliable information about its relatives.
- Grandala, Grandala coelicolor
- Mountain Bluebird, Sialia currucoides
- Western Bluebird, Sialia mexicana
- Eastern Bluebird, Sialia sialis
- Boulder Chat, Pinarornis plumosus
- White-tailed Ant-Thrush, Neocossyphus poensis
- Red-tailed Ant-Thrush, Neocossyphus rufus
- Finsch's Rufous-Thrush, Stizorhina finschi
- Fraser's Rufous-Thrush, Stizorhina fraseri
- Townsend's Solitaire, Myadestes townsendi
- Andean Solitaire, Myadestes ralloides
- Black-faced Solitaire, Myadestes melanops
- Varied Solitaire, Myadestes coloratus
- Cuban Solitaire, Myadestes elisabeth
- Rufous-throated Solitaire, Myadestes genibarbis
- Brown-backed Solitaire, Myadestes occidentalis
- Slate-colored Solitaire, Myadestes unicolor
- Kamao, Myadestes myadestinus
- Amaui, Myadestes woahensis
- Olomao, Myadestes lanaiensis
- Omao, Myadestes obscurus
- Puaiohi, Myadestes palmeri
- Sulawesi Thrush, Cataponera turdoides
- Alpine Thrush, Zoothera mollissima
- Himalayan Thrush, Zoothera salimalii
- Sichuan Thrush, Zoothera griseiceps
- Long-tailed Thrush, Zoothera dixoni
- Geomalia, Zoothera heinrichi
- Long-billed Thrush, Zoothera monticola
- Dark-sided Thrush, Zoothera marginata
- Everett's Thrush, Zoothera everetti
- Sunda Thrush, Zoothera andromedae
- White's Thrush, Zoothera aurea
- Scaly Thrush, Zoothera dauma
- Bonin Thrush, Zoothera terrestris
- Nilgiri Thrush, Zoothera neilgherriensis
- Sri Lanka Thrush, Zoothera imbricata
- Amami Thrush, Zoothera major
- White-bellied Thrush, Zoothera margaretae
- Guadalcanal Thrush, Zoothera turipavae
- Bassian Thrush, Zoothera lunulata
- Fawn-breasted Thrush, Zoothera machiki
- Russet-tailed Thrush, Zoothera heinei
- Black-backed Thrush, Zoothera talaseae
- Aztec Thrush, Ridgwayia pinicola
Click for Catharus tree - Varied Thrush, Ixoreus naevius
- Rufous-brown Solitaire, Cichlopsis leucogenys
- Black Solitaire, Entomodestes coracinus
- White-eared Solitaire, Entomodestes leucotis
- Wood Thrush, Hylocichla mustelina
- Black-headed Nightingale-Thrush, Catharus mexicanus
- Orange-billed Nightingale-Thrush, Catharus aurantiirostris
- Spotted Nightingale-Thrush, Catharus dryas
- Slaty-backed Nightingale-Thrush, Catharus fuscater
- Swainson's Thrush, Catharus ustulatus
- Veery, Catharus fuscescens
- Gray-cheeked Thrush, Catharus minimus
- Bicknell's Thrush, Catharus bicknelli
- Hermit Thrush, Catharus guttatus
- Russet Nightingale-Thrush, Catharus occidentalis
- Ruddy-capped Nightingale-Thrush, Catharus frantzii
- Black-billed Nightingale-Thrush, Catharus gracilirostris
- Fruithunter, Chlamydochaera jefferyi
- Purple Cochoa, Cochoa purpurea
- Green Cochoa, Cochoa viridis
- Javan Cochoa, Cochoa azurea
- Sumatran Cochoa, Cochoa beccarii
- Siberian Thrush, Geokichla sibirica
- Pied Thrush, Geokichla wardii
- Black-eared Ground-Thrush, Geokichla camaronensis
- Gray Ground-Thrush, Geokichla princei
- Orange Ground-Thrush, Geokichla gurneyi
- Abyssinian Ground-Thrush, Geokichla piaggiae
- Crossley's Ground-Thrush, Geokichla crossleyi
- Oberlander's Ground-Thrush, Geokichla oberlaenderi
- Spotted Ground-Thrush, Geokichla guttata
- Spot-winged Thrush, Geokichla spiloptera
- Orange-headed Thrush, Geokichla citrina
- Ashy Thrush, Geokichla cinerea
- Red-backed Thrush, Geokichla erythronota
- Red-and-black Thrush, Geokichla mendeni
- Slaty-backed Thrush, Geokichla schistacea
- Buru Thrush, Geokichla dumasi
- Seram Thrush, Geokichla joiceyi
- Orange-sided Thrush, Geokichla peronii
- Chestnut-capped Thrush, Geokichla interpres
- Enggano Thrush, Geokichla leucolaema
- Chestnut-backed Thrush, Geokichla dohertyi
- Chinese Thrush, Otocichla mupinensis
Click for Turdus tree - Ethiopian Thrush, Psophocichla simensis
- Groundscraper Thrush, Psophocichla litsitsirupa
- Mistle Thrush, Turdus viscivorus
- Song Thrush, Turdus philomelos
- Kurrichane Thrush, Turdus libonyana
- Comoros Thrush, Turdus bewsheri
- Chinese Blackbird, Turdus mandarinus
- Bare-eyed Thrush, Turdus tephronotus
- Yemen Thrush, Turdus menachensis
- Somali Thrush, Turdus ludoviciae
- Usambara Thrush, Turdus roehli
- Abyssinian Thrush, Turdus abyssinicus
- Taita Thrush, Turdus helleri
- Karoo Thrush, Turdus smithi
- Olive Thrush, Turdus olivaceus
- African Thrush, Turdus pelios
- Principe Thrush, Turdus xanthorhynchus
- Sao Tome Thrush, Turdus olivaceofuscus
- Redwing, Turdus iliacus
- Eurasian Blackbird / Common Blackbird, Turdus merula
- Sulawesi Thrush, Turdus turdoides
- Fieldfare, Turdus pilaris
- Chestnut Thrush, Turdus rubrocanus
- White-collared Blackbird, Turdus albocinctus
- Ring Ouzel, Turdus torquatus
- Black-throated Thrush, Turdus atrogularis
- Red-throated Thrush, Turdus ruficollis
- Dusky Thrush, Turdus eunomus
- Naumann's Thrush, Turdus naumanni
- Taiwan Thrush, Turdus niveiceps
- Gray-winged Blackbird, Turdus boulboul
- Indian Blackbird, Turdus simillimus
- Tickell's Thrush, Turdus unicolor
- Black-breasted Thrush, Turdus dissimilis
- Gray-backed Thrush, Turdus hortulorum
- Japanese Thrush, Turdus cardis
- Tibetan Blackbird, Turdus maximus
- Kessler's Thrush, Turdus kessleri
- Brown-headed Thrush, Turdus chrysolaus
- Izu Thrush, Turdus celaenops
- Pale Thrush, Turdus pallidus
- Eyebrowed Thrush, Turdus obscurus
- Gray-sided Thrush, Turdus feae
- Mindoro Island Thrush, Turdus mindorensis
- Luzon Island Thrush, Turdus thomassoni
- Mindanao Island Thrush, Turdus nigrorum
- Sundaic Island Thrush, Turdus javanicus
- Wallacean Island Thrush, Turdus schlegelii
- Christmas Island Thrush, Turdus erythropleurus
- Moluccan Island Thrush, Turdus deningeri
- Papuan Island Thrush, Turdus papuensis
- Bismarck Island Thrush, Turdus heinrothi
- Bougainville Island Thrush, Turdus bougainvillei
- Solomons Island Thrush, Turdus kulambangrae
- Vanikoro Island Thrush, Turdus vanikorensis
- Banks Island Thrush, Turdus whitneyi
- White-headed Island Thrush, Turdus pritzbueri
- New Caledonian Island Thrush, Turdus xanthopus
- Tasman Sea Island Thrush, Turdus poliocephalus
- Samoan Island Thrush, Turdus samoensis
- Fiji Island Thrush, Turdus ruficeps
- Red-legged Thrush, Turdus plumbeus
- †Grand Cayman Thrush, Turdus ravidus
- White-chinned Thrush, Turdus aurantius
- American Robin, Turdus migratorius
- Rufous-collared Robin / Rufous-collared Thrush, Turdus rufitorques
- Black Thrush, Turdus infuscatus
- Sooty Thrush, Turdus nigrescens
- Mountain Thrush, Turdus plebejus
- White-eyed Thrush, Turdus jamaicensis
- La Selle Thrush, Turdus swalesi
- Forest Thrush, Turdus lherminieri
- Pale-eyed Thrush, Turdus leucops
- Chestnut-bellied Thrush, Turdus fulviventris
- Plumbeous-backed Thrush, Turdus reevei
- Chiguanco Thrush, Turdus chiguanco
- Andean Slaty Thrush, Turdus nigriceps
- Glossy-black Thrush, Turdus serranus
- Great Thrush, Turdus fuscater
- Black-hooded Thrush, Turdus olivater
- Austral Thrush, Turdus falcklandii
- Tristan Thrush, Turdus eremita
- Creamy-bellied Thrush, Turdus amaurochalinus
- Black-billed Thrush, Turdus ignobilis
- Maranon Thrush, Turdus maranonicus
- Campina Thrush, Turdus arthuri
- Yellow-legged Thrush, Turdus flavipes
- White-necked Thrush, Turdus albicollis
- Dagua Thrush, Turdus daguae
- White-throated Thrush, Turdus assimilis
- Pale-vented Thrush, Turdus obsoletus
- Rufous-backed Robin / Rufous-backed Thrush, Turdus rufopalliatus
- Lawrence's Thrush, Turdus lawrencii
- Pantepui Thrush, Turdus murinus
- Pale-breasted Thrush, Turdus leucomelas
- Cocoa Thrush, Turdus fumigatus
- Hauxwell's Thrush, Turdus hauxwelli
- Blacksmith Thrush, Turdus subalaris
- Rufous-bellied Thrush, Turdus rufiventris
- Unicolored Thrush, Turdus haplochrous
- Spectacled Thrush, Turdus nudigenis
- Varzea Thrush, Turdus sanchezorum
- Clay-colored Thrush, Turdus grayi
- Ecuadorian Thrush, Turdus maculirostris
Muscicapidae: Old World Flycatchers, Chats Fleming, 1822
59 genera, 335 species HBW-10 & 11
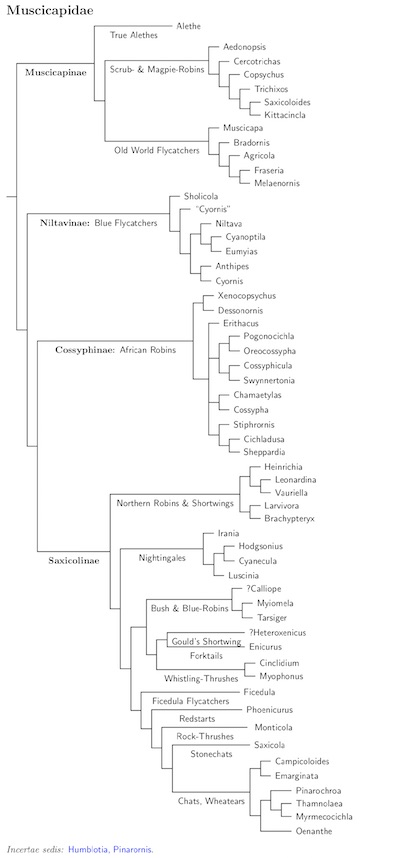 The Muscicapidae have been a very troublesome family for the TiF list. They have
undergone several reorganizations as new data become available. The lastest is
version 2.60 which incorporates Sangster et. al. (2010). I'm happy to say that
their results are generally congruent with Zuccon and Ericson (2010c), so that
most of the large-scale features of the 2.54 reorganization are retained. Even
the 2.54 changes were less extensive than the 2.10 reorganization, and it seems
were are converging on the true phylogenetic tree. Most of the differences that
exist between Sangster et al. (2010) and Zuccon and Ericson (2010c) are within
the major clades, not between them.
The Muscicapidae have been a very troublesome family for the TiF list. They have
undergone several reorganizations as new data become available. The lastest is
version 2.60 which incorporates Sangster et. al. (2010). I'm happy to say that
their results are generally congruent with Zuccon and Ericson (2010c), so that
most of the large-scale features of the 2.54 reorganization are retained. Even
the 2.54 changes were less extensive than the 2.10 reorganization, and it seems
were are converging on the true phylogenetic tree. Most of the differences that
exist between Sangster et al. (2010) and Zuccon and Ericson (2010c) are within
the major clades, not between them.
Both Sangster et al. (2010) and Zuccon and Ericson (2010c) are multi-gene analyses with large samples of species. Both use about the same amount of DNA from 4 genes, with a 2 gene overlap between the papers. Zuccon and Ericson sampled 66 species of Muscicapidae, while Sangster et al. examined 124 species of Muscicapidae.
There are still problems because some generic boundaries need to be redrawn (e.g., see Seki, 2006), and there are issues all the way down to the species level (e.g., Stonechats). The starting point is now Sangster et al. (2010), supplemented by Zuccon and Ericson (2010c). To this I have added information from a number of additional papers: Beresford (2003), Cibois and Cracraft (2004), Illera et al. (2008), Lei et al. (2007), Moyle et al. (2005), Outlaw and Voelker (2006), Outlaw et al. (2007), Outlaw et al. (2010), Pan et al. (2006), Seki (2006), Sheldon et al. (2009), Voelker (2010), Voelker and Spellman (2004), Wink et al. (2002), and Zuccon and Ericson (2010a). From these, I came up with the tree on the right. Earlier versions relied on Jønsson and Fjeldså's supertree (2006a), but it has little direct impact on this version.
The relationships amoung the Turdidae (traditionally including robins, chats, and thrushes) and the Muscicapidae (flycatchers) have long been confusing. Sibley and Monroe grouped them into a large Muscicapidae family with the thrushes in Turdinae, flycatchers as tribe Muscicapini (in subfamily Muscicapinae), and robins and chats in tribe Saxicolini (also Muscicapinae). A similar arrangement is followed here, with families Turdidae and Muscicapidae, the latter divided into Muscicapinae and Saxicolinae. However, some of the genera invovled had hopped from one group to another. This is pretty obvious when we look at the Muscicapinae.
Muscicapinae Fleming, 1822
One genus from Sibley and Monroe's Turdinae (Alethe) is basal. The situation is actually a little more complex than it seems as Beresford (2003) split Alethe, with part (Chamaetylas) moving to Saxicolinae. None remained in Turdidae. There is a conflict here between Sangster et al. (2010) and Zuccon and Ericson (2010c). I've used the Zuccon and Ericson solution (placing Alethe basally in Muscicapinae rather than including it in Copsychini because they sampled both species of Alethe while Sangster et al. sampled only one. It is well-known that sampling two taxa from a genus is more likely to give reliable results.
The remainder of the subfamily splits into two groups, Copsychini and Muscicapini.
I'm not at all sure where Humblotia fits, so I've placed it at the end with the other incertae sedis species, even though it is believed to be closely related to Muscicapa.
Alethini: True Alethes Informal?
- Fire-crested Alethe, Alethe castanea
- White-tailed Alethe, Alethe diademata
Copsychini: Scrub-Robins, Magpie-Robins, Shama
Copsychini contains five genera that Sibley and Monroe put in Saxicolini (Saxicoloides, Trichixos, Copsychus, Cercotrichas, and Erythropygia). It turns out that some of the Cercotrichas form a basal clade. These have been separated in the genus Aedonopsis (Sharpe 1883, type signata). This has been regarded as preoccupied by Aedonopsis Rey 1872, which is an incorrect spelling of Aedonops Brehm 1863. As such, it is not available to preoccupy anything. See Raty, BirdForum 2018. As a result, we don't use the replacement name Tychaedon (Richmond 1917, type signata). The remaining Cercotrichas (Cercotrichas Boie 1831 has priority over Erythropygia Smith 1836) are sister to the shamas and magpie-robins.
There is a little more uncertainty about the relationship of the other shamas to Rufous-tailed Shama (Trichixos). Sangster et al. (2010), Voelker et al. (2014), Zuccon and Ericson (2010c) find Trichixos groups with the other shamas and Saxicoloides, but Lim et al. (2010b) put the Rufous-tailed Shama closer to the magpie-robins. I've grouped it with the shamas and Indian Robin (Saxicoloides), placing the magpie-robins in Copsychus (Wagler 1827). The shamas are separated in Kittacincla (Gould 1831, type malabaricus) for the shamas. I retain Saxicoloides (Lesson 1831), Trichixos (Lesson 1839), and Kittacincla (Gould 1836) as separate genera due to their distinct appearance. However, given the genetic distances involved, it would not be unreasonable to combine them into Saxicoloides or even combine all three with Copsychus.
The Maputaland Scrub-Robin, Aedonopsis tongensis, has been split from the Brown Scrub-Robin, Aedonopsis signata. See Ribeiro et al. (2014). Also, the Philippine Magpie-Robin, Copsychus mindanensis, has been split from Oriental Magpie-Robin, Copsychus saularis, by Sheldon et al. (2009). It appears to be basal among the Magpie-Robins.
Copsychini: Scrub-Robins, Magpie-Robins, Shamas Sundevall, 1872
- Karoo Scrub-Robin, Aedonopsis coryphoeus
Click for Copsychini tree - Brown Scrub-Robin, Aedonopsis signata
- Maputaland Scrub-Robin, Aedonopsis tongensis
- Forest Scrub-Robin, Aedonopsis leucosticta
- Bearded Scrub-Robin, Aedonopsis quadrivirgata
- Miombo Scrub-Robin, Aedonopsis barbata
- Kalahari Scrub-Robin, Cercotrichas paena
- Rufous-tailed Scrub-Robin, Cercotrichas galactotes
- Black Scrub-Robin, Cercotrichas podobe
- Brown-backed Scrub-Robin, Cercotrichas hartlaubi
- White-browed Scrub-Robin, Cercotrichas leucophrys
- Philippine Magpie-Robin, Copsychus mindanensis
- Madagascan Magpie-Robin, Copsychus albospecularis
- Seychelles Magpie-Robin, Copsychus sechellarum
- Oriental Magpie-Robin, Copsychus saularis
- Indian Robin, Saxicoloides fulicatus
- Rufous-tailed Shama, Trichixos pyrropygus
- White-browed Shama, Kittacincla luzoniensis
- White-rumped Shama, Kittacincla malabarica
- Andaman Shama, Kittacincla albiventris
- White-vented Shama, Kittacincla nigra
- Black Shama, Kittacincla cebuensis
Muscicapini: Old World Flycatchers
We now turn to Muscicapini. Using the terminology of Dickinson (2003), Muscicapini is restricted to Empidornis, Fraseria, Melaenornis, Muscicapa, and Myioparus. The more recent H&M-4v2 (Dickinson and Christidis, 2014) recognizes Muscicapini and divides it into Muscicapa, Myioparus, Fraseria, Bradornis, Melaenornis, Empidornis, Sigelus, and Humblotia.
Neither Zuccon and Ericson (2010c) nor Sangster et al (2010) sampled enough species to clarify the structure of Muscicapini. That has changed with Voelker et al. (2016), who sampled most of the Muscicapini.
There are various ways the tribe can be divided into genera. Voelker et al. seem to prefer more rather than fewer. However, their molecular clock analysis suggests that the entire tribe is fairly closely related (within 7-8 million years, 6-7 million without the basal species). Some people would respond to that by putting them all in one genus! I've taken a middle course.
The Rusty-tailed Flycatcher, Muscicapa ruficauda, has been moved to Ficedula. This is based on the phylogenetic trees associated with Price et al. (2014) and Raty's analysis on BirdForum. See also Hooper et al. (2016)
Next is a reduced genus Muscicapa. As this group includes the type species (striata), it retains the name Muscicapa.
The correct scientific name of the Asian Brown Flycatcher appears to be Muscicapa latirostris not Muscicapa dauurica (Mlíkovský, 2012). Further, the Brown-streaked Flycatcher, Muscicapa williamsoni (including umbrosa), has been split from the Asian Brown Flycatcher (Rheindt and Eaton, 2012). For the present, I'm treating Muscicapa sodhii (Harris et al., 2015) as a subspecies of the Asian Brown Flycatcher, Muscicapa latirostris. The available genetic data place it close to the subspecies siamensis, closer than either is to the nominate.
Based on Pons et al. (2016), the Mediterranean Flycatcher, Muscicapa tyrrhenica (including balearica) has been split from Spotted Flycatcher, Muscicapa striata. See Viganó and Corso (2015) concerning identification of balearica. There is some possibility of a further split into monotypic species, which would probably be named Balearic and Tyrrhenian Flycatchers.
Next in line is Bradornis (Smith 1847, type mariquensis). Two species, Pale Flycatcher, formerly ``Bradornis'' pallidus, and Chat Flycatcher, formerly Bradornis infuscatus have been moved to a separate genus, Agricola (Bonaparte 1854, type infuscatus) This has priority over Sericolius (Bonaparte 1855, type pallidus). This is the next branch. There is a name complication here as both Sooty Flycatcher and Chat Flycatcher are named infuscatus. The Chat Flycatcher was originally called Saxicola infuscata by A. Smith in 1839, while the Sooty Flycatcher was dubbed Batails infuscatus by Cassin in 1855. The Chat Flycatcher gets the name infuscatus back as it is now in a separate genus.
Bradornis also gains 5 species compared to H&M-4. I had previously transferred Ussher's Flycatcher, Bradornis ussheri, and Dusky-blue Flycatcher, Bradornis comitata from Muscicapa. Voelker's analysis also adds Boehm's Flycatcher, formerly Muscicapa boehmi, and Dusky-blue Flycatcher, formerly Muscicapa comitata, to the group.
The fifth species is the Sooty Flycatcher, sometimes called Muscicapa infuscata. It was originally named Artomyias fuliginosa by J. and E. Verreaux in 1855. Once upon a time this conflicted with Muscicapa fuliginosa Sparrman 1787, but that species is now in Rhipidura, far away from any of the Muscicapidae. If we retained the Sooty Flycatcher in Muscicapa, there'd be another fuliginosa conflict, as the name was also applied by Hodgson in 1845 to what is now Muscicapa sibirica cacabata. The name cacabata was coined by Penard in 1919 in order to avoid a conflict with the Rhipidura species. Since I'm treating the Sooty Flycatcher as part of Bradornis, which contains neither of those species, it is safe to call the Sooty Flycatcher Bradornis fuliginosa.
There are two clades after Agricola. The oldest names available for them are Fraseria (Bonaparte 1854, type ocreata and Melaenornis (G.R. Gray 1840, type edolioides), respectively.
In H&M-4, Fraseria consisted of two species. Now it has grown to 7. The latest additions are White-browed Forest-Flycatcher, formerly Melaenornis cinerascens (already Fraseria in H&M-4) and Tessmann's Flycatcher, formerly Muscicapa tessmanni. I had previously added the two Myioparus based on Zuccon and Ericson (2010c). I had also previously added 3 former Muscicapa Ashy Flycatcher, Fraseria caerulescens (see Sangster et al, 2010; Zuccon and Ericson, 2010c); Olivaceous Flycatcher, Fraseria olivascens (see Sangster et al., 2010); and Chapin's Flycatcher, Fraseria lendu (it may belong near olivascens, but I've seen no genetic evidence for this).
That brings us to Melaenornis. It had earlier absorbed the monotypic genera Sigelus and Empidornis based on Zuccon and Ericson (2010c). Now, it surprisingly absorbs Namibornis, the Herero Chat. The chat had been difficult to place and some authors put it in Turdidae. I had previously left it Incertae sedis in Muscicapidae. Voelker et al. (2016) found it belongs in Melaenornis. The remaining Melaenornis agrees with H&M-4.
Muscicapini: Old World Flycatchers Fleming, 1822
- Gray-streaked Flycatcher, Muscicapa griseisticta
Click for Muscicapini tree - Dark-sided Flycatcher, Muscicapa sibirica
- Ferruginous Flycatcher, Muscicapa ferruginea
- Brown-breasted Flycatcher, Muscicapa muttui
- Asian Brown Flycatcher, Muscicapa latirostris
- Brown-streaked Flycatcher, Muscicapa williamsoni
- Ashy-breasted Flycatcher, Muscicapa randi
- Sumba Brown Flycatcher, Muscicapa segregata
- Little Gray Flycatcher, Muscicapa epulata
- African Dusky Flycatcher, Muscicapa adusta
- Yellow-footed Flycatcher, Muscicapa sethsmithi
- Spotted Flycatcher, Muscicapa striata
- Mediterranean Flycatcher, Muscicapa tyrrhenica
- Gambaga Flycatcher, Muscicapa gambagae
- Swamp Flycatcher, Muscicapa aquatica
- Cassin's Flycatcher, Muscicapa cassini
- Boehm's Flycatcher, Bradornis boehmi
- Sooty Flycatcher, Bradornis fuliginosus
- Ussher's Flycatcher, Bradornis ussheri
- Dusky-blue Flycatcher, Bradornis comitata
- African Gray Flycatcher, Bradornis microrhynchus
- Marico Flycatcher, Bradornis mariquensis
- Pale Flycatcher, Agricola pallidus
- Chat Flycatcher, Agricola infuscatus
- White-browed Forest-Flycatcher, Fraseria cinerascens
- Fraser's Forest-Flycatcher, Fraseria ocreata
- Gray Tit-Flycatcher, Fraseria plumbea
- Gray-throated Tit-Flycatcher, Fraseria griseigularis
- Ashy Flycatcher, Fraseria caerulescens
- Tessmann's Flycatcher, Fraseria tessmanni
- Olivaceous Flycatcher, Fraseria olivascens
- Chapin's Flycatcher, Fraseria lendu
- Fiscal Flycatcher, Melaenornis silens
- Herero Chat, Melaenornis herero
- Silverbird, Melaenornis semipartitus
- Abyssinian Slaty-Flycatcher, Melaenornis chocolatinus
- Angolan Slaty-Flycatcher, Melaenornis brunneus
- White-eyed Slaty-Flycatcher, Melaenornis fischeri
- Yellow-eyed Black-Flycatcher, Melaenornis ardesiacus
- Nimba Flycatcher, Melaenornis annamarulae
- Northern Black-Flycatcher, Melaenornis edolioides
- Southern Black-Flycatcher, Melaenornis pammelaina
Niltavinae: Blue Flycatchers Sangster et al., 2010
Both Zuccon and Ericson (2010c) and Sangster et al. (2010) found a clade containing the blue flycatchers. Sangster et al. recommended treating it as a subfamily and proposed the name Niltavinae.
Sangster et al. considered several species of Rhinomyias and discovered it was not a natural group (there seem to be at least 4 pieces). Much of Rhinomyias seems to be embedded in Cyornis. Those taxa are merged here. It's a little surprising to include the drab Rhinomyias in Cyornis. Further analysis should help us make better sense of this. The Streak-breasted Jungle-Flycatcher, Rhinomyias additus, has been transferred to Eumyias and several other members of Rhinomyias have been transferred to Vauriella in Saxicolinae. Surprisingly, Leonardina, thought to be a babbler, is closely related to Vauriella (Oliveros et al., 2012). Indeed, it might even be better to sink it in Vauriella.
Based on Robin et al. (2017), the Nilgiri Blue-Robin, Myiomela major, and White-bellied Blue-Robin, Myiomela albiventris, have been separated in the new genus Sholicola. I have changed the primary name to Sholakili to reflect this (note that Blue-Flycatcher would lead to a name conflict). Also, I have accepted the split of Ashambu Sholakili, Sholicola ashambuensis, from White-bellied Sholakili, Sholicola albiventris (Robin et al., 2017).
The new genus Sholicola is basal in this group, likely followed by the White-tailed Flycatcher (Robin et al., 2017, have it in a slightly different position). Whether it is alone or whether other species are with it is not yet clear. We use the temporary genus “Cyornis” until this situation is straightened out.
The remainder of Niltavinae breaks into two parts. The first consists of Cyanoptila, Eumyias, and Niltava. Although Lei et al. (2007), using less data, considered this group part of Muscicapinae (as did Sibley and Monroe), Voelker and Spellman (2004), Sangster et al. (2010), and Zuccon and Ericson (2010c) found the contrary. The second clade includes Anthipes (previously removed from Ficedula by Outlaw and Voelker, 2006a), the Cyornis blue-flycatchers and jungle-flycatchers (transferred from Rhinomyias).
Zappey's Flycatcher, Cyanoptila cumatilis, has been split from the Blue-and-white Flycatcher, Cyanoptila cyanomelana (Leader and Carey, 2012).
The Chinese Blue-Flycatcher, Cyornis glaucicomans, has been split from Blue-throated Blue-Flycatcher, Cyornis rubeculoides. Note that the subspecies klossi seems to belong to the Hainan Blue-Flycatcher, Cyornis hainanus. See Zhang et al. (2016).
- Nilgiri Blue-Flycatcher / Nilgiri Blue-Robin, Sholicola major
- White-bellied Blue-Flycatcher / White-bellied Blue-Robin, Sholicola albiventris
- Ashambu Blue-Flycatcher / Ashambu Blue-Robin, Sholicola ashambuensis
- White-tailed Flycatcher, "Cyornis" concretus
- Rufous-bellied Niltava, Niltava sundara
- Small Niltava, Niltava macgrigoriae
- Rufous-vented Niltava, Niltava sumatrana
- Vivid Niltava, Niltava vivida
- Fujian Niltava, Niltava davidi
- Large Niltava, Niltava grandis
- Zappey's Flycatcher, Cyanoptila cumatilis
- Blue-and-white Flycatcher, Cyanoptila cyanomelana
- Nilgiri Flycatcher, Eumyias albicaudatus
- Indigo Flycatcher, Eumyias indigo
- Dull-blue Flycatcher, Eumyias sordidus
- Verditer Flycatcher, Eumyias thalassinus
- Turquoise Flycatcher, Eumyias panayensis
- Streak-breasted Jungle-Flycatcher, Eumyias additus
- White-gorgeted Flycatcher, Anthipes monileger
- Rufous-browed Flycatcher, Anthipes solitaris
- Russet-backed Jungle-Flycatcher, Cyornis oscillans
- Brown-chested Jungle-Flycatcher, Cyornis brunneatus
- Nicobar Jungle Flycatcher, Cyornis nicobaricus
- Fulvous-chested Jungle-Flycatcher, Cyornis olivaceus
- Gray-chested Jungle-Flycatcher, Cyornis umbratilis
- Rufous-tailed Jungle-Flycatcher, Cyornis ruficauda
- Henna-tailed Jungle-Flycatcher, Cyornis colonus
- Pale Blue-Flycatcher, Cyornis unicolor
- Rueck's Blue-Flycatcher, Cyornis ruckii
- Blue-breasted Blue-Flycatcher, Cyornis herioti
- White-bellied Blue-Flycatcher, Cyornis pallipes
- Pale-chinned Blue-Flycatcher, Cyornis poliogenys
- Hill Blue-Flycatcher, Cyornis banyumas
- Large Blue-Flycatcher, Cyornis magnirostris
- Palawan Blue-Flycatcher, Cyornis lemprieri
- Tickell's Blue-Flycatcher, Cyornis tickelliae
- Sunda Blue-Flycatcher, Cyornis caerulatus
- Bornean Blue-Flycatcher, Cyornis superbus
- Chinese Blue-Flycatcher, Cyornis glaucicomans
- Hainan Blue-Flycatcher, Cyornis hainanus
- Blue-throated Blue-Flycatcher, Cyornis rubeculoides
- Malaysian Blue-Flycatcher, Cyornis turcosus
- Mangrove Blue-Flycatcher, Cyornis rufigastra
- Sulawesi Blue-Flycatcher, Cyornis omissus
- Timor Blue-Flycatcher, Cyornis hyacinthinus
- Blue-fronted Blue-Flycatcher, Cyornis hoevelli
- Matinan Blue-Flycatcher, Cyornis sanfordi
Cossyphinae: African Robins Vigors, 1825
The African Robins, Cossyphinae, are the most problematic part of the Muscicapidae. There are many studies concerning them: Roy et al. (2000), Beresford (2003), and Voelker et al. (2010a) focus on the group, and it is an important component of the large-scale analyses of Zuccon and Ericson (2010c) and Sangster et al. (2010). Moreover, a number of other papers have focused on individual species or small groups of species. When all of this is put together, the result is confusion. There is no argreement on how the genera are arranged, and there are still questions about which species belong to which genera and about which species are actually species. The only really consistent point is the list of taxa that belong to Cossyphinae.
With that in mind, I'm now using Sangster et al. (2010) as the basic framework. It is not really satisfactory as it doesn't provide a lot of resolution. Moreover, comparison with Zuccon and Ericson (2010c) makes one wonder if even the terminal clades are right.
Even the name of this group is a problem. Sangster et al. (2010) refer to this subfamily as Erithacinae, citing G.R. Gray 1846 (priority seems to date to 1831 under a different name). Still, the name Cossyphinae dates to Vigors 1825 (as Cossyphina), and should have priority.
The sequence starts with Xenocopsychus and Dessonornis (A. Smith 1836, type humeralis). Although Sangster et al. only included archeri and humeralis, Voelker et al. (2010a) also grouped caffra and anomala with archeri. Using substantially less data, Beresford (2003) grouped caffra and archeri, but excluded anomala. Including humeralis allows us to use the name Dessonornis. Finally, Xenocopsychus has been thought to be close to humeralis.
Voelker et al. (2010a) advocated use of the name Callene. The name Callene is due to Blyth (1847). He introduced it as a substitute for Cinclidium. He felt this was necessary as Cinclidium was already in use in botany for a type of moss. These days, the same genus name can and is used in both botany and zoology without conflict, and Cinclidium has regained its name. Although it subsequently became a larger genus (and later shrank), when Blyth proposed Callene, Cinclidium contained only one species, Cinclidium frontale. That means the type of Callene is Cinclidium frontale. If humeralis were not there, the oldest name having any of these as type species is Caffronis (Roberts, 1922; type C. caffra).
The remainder is organized as a four-fold polytomy, which is a way of say the data is pretty inconclusive. One piece of the polytomy is the only non-African species in the group, the Eurasian Robin.
A second piced consists of Pogonocichla, Oreocossypha, Cossyphicula, and Swynnertonia. Beresford found evidence that all three form a clade. It might be reasonable to place these species in the same genus, but it's probably better to wait for more evidence.
Beresford (2003) found that several members of Alethe were not closely related to the other Alethe. In fact, it turns out they do not belong in Muscicapinae at all. Rather, they belong with the forest-robins and take the genus name Chamaetylas, part of Cossyphinae. Sangster et al. (2010) group them with a somewhat restructered Cossypha.
Beresford also found evidence that both Sheppardia and Cossypha were paraphyletic. Recently, Voelker et al. (2010a) focused on Sheppardia and the related species in Cossypha. Their analysis is the basis of the arrangement here. The Gray-winged Robin-Chat, formerly Cossypha polioptera, has been transferred to Sheppardia. Three other members of Cossypha, caffra, anomala, and archeri, have been placed in a separate genus, Dessonornis.
The last of the 4 clades includes Stiphrornis, Cichladusa, and the revised Sheppardia. Only Voelker et al. (2010a) analyzed Cichladusa, finding that it is near Sheppardia, but not so close to Stiphrornis.
On the IOC list, the various Stiphrornis forest-robins are considered a single species, the Forest Robin. Cornell considers them as three species: Olive-backed Forest Robin, Stiphrornis pyrrholaemus, Orange-breasted Forest Robin, Stiphrornis erythrothorax, and Yellow-breasted Forest Robin, Stiphrornis mabirae. I had previously considered them to be 5 species based on Beresford and Cracraft (1999) and Schmidt et al. (2008).
Well, time has progressed, and two more papers have appeared, Voelker et al. (2017) and Dongmo et al. (2019), and I still see 5 clades of what appear to be species-level taxa, with both the Orange-breasted Forest Robin, Stiphrornis erythrothorax, and Yellow-breasted Forest Robin, Stiphrornis mabirae, representing two species. However, more data has made the divisions seem smaller, and it could well end up as 3 or 4. Right now it looks like 5, and I continue to go with that. We can allocate the subspecies better and Birds of the World even has relevant range maps and illustrations.
Here's my old arrangement:
- Eastern Forest-Robin, Stiphrornis xanthogaster
- Sangha Forest-Robin, Stiphrornis sanghensis
- Gabon Forest-Robin, Stiphrornis gabonensis
- Western Forest-Robin, Stiphrornis erythrothorax
- Olive-backed Forest-Robin, Stiphrornis pyrrholaemus
and here's the new:
- Olive-backed Forest-Robin, Stiphrornis pyrrholaemus (monotypic)
- Western Orange-breasted Forest-Robin, Stiphrornis erythrothorax (monotypic)
- Eastern Orange-breasted Forest-Robin, Stiphrornis gabonensis, including dahomeyensis and inexpectatus.
- Yellow-bellied Forest-Robin, Stiphrornis xanthogaster, including sanghensis and rudderi
- Yellow-breasted Forest-Robin, Stiphrornis mabirae
The subspecies xanthogaster is a problem as the birds further east appear different from those to the west. Fortunately, the name mabirae (Jackson 1910), type Mariba Forest, Cental African Republic, is available for eastern birds, leaving xanthogaster (Sharpe, 1903), with type from the Dja River in SE Cameroon available for those in the western Congo basin. Unfortunately, it appears that all of the sampled `xanthogaster' are actually mabirae. I'm guessing that the true xanthogaster would group with sanghensis and rudderi.
- Angolan Cave Chat, Xenocopsychus ansorgei
- Cape Robin-Chat, Dessonornis caffra
- Olive-flanked Robin-Chat / Olive-flanked Ground-Robin, Dessonornis anomala
- Archer's Robin-Chat / Archer's Ground-Robin, Dessonornis archeri
- White-throated Robin-Chat, Dessonornis humeralis
- European Robin, Erithacus rubecula
- White-starred Robin, Pogonocichla stellata
- Mountain Robin-Chat, Oreocossypha isabellae
- White-bellied Robin-Chat, Cossyphicula roberti
- Swynnerton's Robin, Swynnertonia swynnertoni
- Red-throated Alethe, Chamaetylas poliophrys
- Brown-chested Alethe, Chamaetylas poliocephala
- White-chested Alethe, Chamaetylas fuelleborni
- Thyolo Alethe, Chamaetylas choloensis
- Blue-shouldered Robin-Chat, Cossypha cyanocampter
- Rueppell's Robin-Chat, Cossypha semirufa
- White-browed Robin-Chat, Cossypha heuglini
- Red-capped Robin-Chat, Cossypha natalensis
- Chorister Robin-Chat, Cossypha dichroa
- White-headed Robin-Chat, Cossypha heinrichi
- Snowy-crowned Robin-Chat, Cossypha niveicapilla
- White-crowned Robin-Chat, Cossypha albicapillus
- Olive-backed Forest-Robin, Stiphrornis pyrrholaemus
- Western Orange-breasted Forest-Robin, Stiphrornis erythrothorax
- Eastern Orange-breasted Forest-Robin, Stiphrornis gabonensis
- Yellow-bellied Forest-Robin, Stiphrornis xanthogaster
- Yellow-breasted Forest-Robin, Stiphrornis mabirae
- Collared Palm-Thrush, Cichladusa arquata
- Rufous-tailed Palm-Thrush, Cichladusa ruficauda
- Spotted Palm-Thrush, Cichladusa guttata
- Usambara Akalat, Sheppardia montana
- Iringa Akalat, Sheppardia lowei
- Rubeho Akalat, Sheppardia aurantiithorax
- Gabela Akalat, Sheppardia gabela
- Equatorial Akalat, Sheppardia aequatorialis
- Lowland Akalat, Sheppardia cyornithopsis
- East Coast Akalat, Sheppardia gunningi
- Sharpe's Akalat, Sheppardia sharpei
- Gray-winged Robin-Chat, Sheppardia polioptera
- Bocage's Akalat, Sheppardia bocagei
Saxicolinae: Robins, Chats, Wheatears Vigors, 1825
Northern Robins and Shortwings
Another important discrepancy between Zuccon and Ericson (2010c) and Sangster et al. (2010) is the basal group in Saxicolinae. Sangster et al. included the White-browed Jungle-Flycatcher, Rhinomyias insignis. in their analysis. It grouped with a former member of the Turdidae, the Great Shortwing, Heinrichia calligyna. The two were pulled into a group with more former Turdidae, Brachypterx, and some of the northern robins. Of course, most of the Rhinomyias are left in the Muscicapinae with Cyornis. In 1980, Wouters had suggested that insignis and several other Rhinomyias be placed in a new genus, Vauriella. I follow that here.
The former robins were variously placed in Luscinia and Erithacus. However, neither of those type species is in this group and they take the name Larvivora (Hodgson 1837, type cyane). Sangster et al. (2010) suggested that the the Rufous-headed Robin, Larvivora ruficeps, belongs here. Larvivora is sister to the Brachypteryx shortwings. More recently, Zhao et al. (2016a) analyzed DNA from all 6 Lavivora species and confirmed that the Rufous-headed Robin belongs in the Larvivora clade, and that it is sister to the Rufous-tailed Robin, Larvivora sibilans.
Based on Kyriazis et al. (2018) and the HBW-Checklist (del Hoyo and Collar, 2016), I've split the White-browed Shortwing into:
- Himalayan Shortwing, Brachypteryx cruralis
- Chinese Shortwing, Brachypteryx sinensis
- Taiwan Shortwing, Brachypteryx goodfellowi
- Sumatran Shortwing, Brachypteryx saturata
- Javan Shortwing, Brachypteryx montana
- Flores Shortwing, Brachypteryx floris
- Bornean Shortwing, Brachypteryx erythrogyna
- Philippine Shortwing, Brachypteryx poliogyna
- Mt. Apo Shortwing, Brachypteryx mindanensis
The Lesser Shortwing, Brachypteryx leucophris, is also embedded within this clade.
Nightingales
This group is followed by the nightingales and allies. Here the White-throated Robin, Irania gutturalis, is basal. Then we have the White-bellied Redstart, Hodgsonius phaenicuroides and the Bluethroat on one side, and the two nightingales on the other. I've put the Bluethroat in its own genus, Cyanecula (Brehm 1828).
Bush-Robins and Blue-Robins, Forktails and Whistling-Thrushes
The next clade is somewhat larger, containing two main groups. The first is another collection of former Luscinia, these placed in Calliope (Gould 1836, type calliope). These are sister to the Myiomela blue-robins and Tarsiger bush-robins. The other half of the clade includes the Myophonus whistling-thrushes (former Turdidae), Gould's Shortwing (Heteroxenicus), and the Enicurus forktails (former Muscicapinae). The forktails have been studied by Moyle et al. (2005), and the arrangement here draws on that work.
The position of Gould's Shortwing (formerly in Brachypteryx), is based on Price et al. (2014). Note that Robin et al. (2017) put it in a different place in the tree.
There is some question about whether Calliope belongs here. Price et al. (2014) and Robin et al. (2017) in two other locations.
The White-tailed Rubythroat, Calliope pectoralis, has been split into Himayalan Rubythroat, Calliope pectoralis, and Chinese Rubythroat, Calliope tschebaiewi, based on Liu et al. (2016).
After this, several genera branch off one at a time: the Ficedula flycatchers, Phoenicurus redstars, Monticola rock-thrushes, and Saxicola chats. The Saxicolinae end with the wheatears and allies.
Ficedula
Outlaw and Voelker (2006a) made a comprehensive study of the Ficedula flycatchers, covering most of the species (a less complete treatment is Lei et al., 2007). As a result, two were removed entirely to Anthipes. Further, Muscicapella has been moved into Ficedula. This means that the Pygmy Flycatcher, Muscicapella hodgsoni, is now Ficedula hodgsoni. This creates a name conflict with the Slaty-backed Flycatcher (the fact that it is hodgsonii, not hodgsoni, makes no difference, ICZN Art. 58). Outlaw and Voelker suggested using Ficedula erithacus. However, Siphia erithacus Jerdon and Blyth, 1861 is preoccupied by Siphia erythaca Blyth, 1847, which is a junior synonym of Ficedula mugimaki. Fortunately, this species has several additional names! The next in line are Erythrosterna sordida Godwin-Austen, 1874, Muscicapa amabilis Deignan, 1947, and Muscicapa erwini Wolters, 1950. There don't seem to be any problems with sordida, so the Slaty-backed Flycatcher becomes Ficedula sordida.
The order within Ficedula primarly follows Moyle et al. (2015), which corrected some problems in Outlaw and Voelker (2006a). Some adjustments have been made based on Hooper et al. (2016) and Dong et al. (2015) has also been consulted. Hooper et al. added some samples from the Black-and-orange Flycatcher, Ficedula nigrorufa. Two splits have been accepted in Ficedula. Særte et al. (2001) showed that the Atlas Flycatcher is a separate species. Moyle et al. (2015) found that the Philippine subspecies of the Snowy-browed Flycatcher, Ficedula hyperythra, are only distantly related to the nominate. Although not all subspecies were sampled, it seems likely that the non-Philippine subspecies form one clade, and the Philippine subspecies another. The Philippine group is recognized as a separate species, Ficedula luzoniensis. The English name presents something of a problem. Philippine Snowy-browed Flycatcher has been suggested, but is a poor choice as they are far from being sister species, and moreover, some of the Philippine subspecies lack the white brow. I now use the IOC name Bundok Flycatcher for Ficedula luzoniensis.
The Rusty-tailed Flycatcher, Muscicapa ruficauda, has been moved to Ficedula. This is based on the phylogenetic trees associated with Price et al. (2014) and Raty's analysis on BirdForum. See also Hooper et al. (2016).
Redstarts
The redstarts are often grouped together. Both Pan et al. (2006) and Voelker (2010) found that Phoenicurus is paraphyletic with respect to Chaimarrornis and Rhyacornis. There are two alternatives, either split the first five as Adelura (Bonaparte 1854, type caeruleocephala), or lump them all into Phoenicurus. Since Chaimarrornis and Rhyacornis have often been considered part of Phoenicurus, and since the genetic distances are relatively small, implying the whole clade is only 5-6 million years old (Voelker, 2010), I've merged them all into Phoenicurus. I've also rearranged them based on Voelker et al. (2015). Pan et al. (2006) found schisticeps belongs in the Adelura group. The position of Moussier's Redstart, Phoenicurus moussieri, has been clarified by Sangster et al. (2010), while that of Przevalski's Redstart, Phoenicurus alaschanicus, has been addressed by Hogner et al. (2012).
Rock-Thrushes
There is some question about whether the Monticola rock-thrushes are sister to the chats and wheatear clade. The evidence on whether they are sister is mixed, and Outlaw et al. (2010) show a different arrangement (without details). The Monticola rock-thrushes do not belong in Turdidae (see Wink et al., 2002; Outlaw et al., 2007). They include the Madagascan Pseudocossyphus rock-thrushes. The former Pseudocossyphus rock-thrushes are sometimes treated as 4 species, but Zuccon and Ericson (2010a) found little genetic distinction between M. sharpei, bensoni, and erythronotus, so I've followed their recommendation and lumped them all into M. sharpei. The other former Pseudocossyphus, M. imerinus is retained as a separate species. A more detailed study by Cruaud et al. (2011) supports this treatment of former Pseudocossyphus.
Zuccon and Ericson (2010a) also found that the White-winged Cliff-Chat belongs in Monticola, not Thamnolaea. Moreover, M. solitarius breaks into two clades which are not each other's closest relatives. The second group consists of the races madoci, philippensis, and pandoo, so they take the name Monticola philippensis. The English name Red-bellied Rock-Thrush has been previously used for philippensis, and I use it here. Nonetheless, it is not a very good name as pandoo, which interbreeds widely with philippensis, is not red-bellied. Perhaps something like “Variable Rock-Thrush” would be better. I've mereged Pretoria Rock-Thrush, Monticola pretoriae into Short-toed Rock-Thrush, Monticola brevipes. Zuccon and Ericson (2010a) found little genetic difference between them. There is reportedly extensive interbreeding.
Chats and Wheatears
The chat/wheatear clade has undergone a fair amount of reorganization. The latest is due to the comprehensive multi-gene analysis of Aliabadian et al. (2012). The clade includes the Buff-streaked Chat, Campicoloides bifasciatus, which Illera et al. (2008) removed from Saxicola. The clade also includes the Mountain Wheatear, which Outlaw et al. (2010) found is not part of Oenanthe. A more complete analysis by Voelker et al. (2012) found it embedded in Myrmecocichla. Both Myrmecocichla and Dromolaea have equal priority, but Voelker et al. refer the Mountain Wheatear to Myrmecocichla. I'm not sure if they need to do more to be first revisers, but if so, Aliabadian et al. (2012), who also Myrmecocichla over Dromolaea are then the first resisers. Either way, Myrmecocichla now has priority.
The Moorland Chat has been moved to Pinarochroa from Cercomela as in Outlaw et al. They also dismembered Cercomela, with the species invovled being distributed between Oenanthe and the revived genus Emarginata (Shelly, 1896, type E. sinuata). Aliabadian et al. (2012) found that the White-fronted Black Chat, sometimes placed in its own genus, Pentholaea, is part of the Oenanthe clade.
Although additional analysis is needed to fully clarify the situation, the Ruaha Chat, Myrmecocichla collaris, has been split from Arnott's Chat, Myrmecocichla arnotti, based on Glen et al. (2011). Both are placed in Myrmecocichla, not Pentholaea (see e.g., Voelker et al., 2012). The arrangement of the various small genera is a compromise between Outlaw et al. (2010), Sangster et al. (2010), and Zuccon and Ericson (2010c).
The arrangement of Oenanthe is based primarily on Outlaw et al. (2010), but Aliabadian et al. (2007) was also consulted concerning species not included by Outlaw et al. The treatment of Saxicola here is based on Wink et al. (2002), Illera et al. (2008), Woog et al. (2008), and Zink et al. (2009). This involves splitting the Common Stonechat into at least seven species: Stejneger's, Siberian, Canary Islands, European, African, Madagascan, and Reunion Stonechats. There may still be additional species hiding in the stonechat complex. For ABA-listers, East Siberian Stonechat is known to occur in the ABA area. The status of West Siberian Stonechat in the ABA area is currently unclear to me.
The “Persian Mourning Wheatear”, Oenanthe persica, is split from Mourning Wheatear Oenanthe lugens, based on Förschler et al. (2010a, 2010b). See also Aliabadian et al. (2012). The English name has gotten some use, but has a problem because there is already a Mourning Wheater, and that name would have to be changed also. I hope a novel name will be proposed for O. persica.
- Great Shortwing, Heinrichia calligyna
- Bagobo Babbler, Leonardina woodi
- Eyebrowed Jungle-Flycatcher, Vauriella gularis
- White-throated Jungle-Flycatcher, Vauriella albigularis
- White-browed Jungle-Flycatcher, Vauriella insignis
- Slaty-backed Jungle-Flycatcher, Vauriella goodfellowi
- Siberian Blue-Robin, Larvivora cyane
- Indian Blue-Robin, Larvivora brunnea
- Rufous-tailed Robin, Larvivora sibilans
- Rufous-headed Robin, Larvivora ruficeps
- Japanese Robin, Larvivora akahige
- Ryukyu Robin, Larvivora komadori
- Rusty-bellied Shortwing, Brachypteryx hyperythra
- Himalayan Shortwing, Brachypteryx cruralis
- Chinese Shortwing, Brachypteryx sinensis
- Taiwan Shortwing, Brachypteryx goodfellowi
- Sumatran Shortwing, Brachypteryx saturata
- Javan Shortwing, Brachypteryx montana
- Flores Shortwing, Brachypteryx floris
- Lesser Shortwing, Brachypteryx leucophris
- Bornean Shortwing, Brachypteryx erythrogyna
- Philippine Shortwing, Brachypteryx poliogyna
- Mt. Apo Shortwing, Brachypteryx mindanensis
- White-throated Robin, Irania gutturalis
- White-bellied Redstart, Hodgsonius phaenicuroides
- Bluethroat, Cyanecula svecica
- Thrush Nightingale, Luscinia luscinia
- Common Nightingale, Luscinia megarhynchos
- Firethroat, Calliope pectardens
- Blackthroat, Calliope obscura
- Siberian Rubythroat, Calliope calliope
- Himayalan Rubythroat, Calliope pectoralis
- Chinese Rubythroat, Calliope tschebaiewi
- White-tailed Robin, Myiomela leucura
- Sunda Robin, Myiomela diana
- Rufous-breasted Bush-Robin, Tarsiger hyperythrus
- Red-flanked Bluetail, Tarsiger cyanurus
- Himalayan Bluetail, Tarsiger rufilatus
- White-browed Bush-Robin, Tarsiger indicus
- Collared Bush-Robin, Tarsiger johnstoniae
- Golden Bush-Robin, Tarsiger chrysaeus
- Gould's Shortwing, Heteroxenicus stellatus
- Little Forktail, Enicurus scouleri
- Slaty-backed Forktail, Enicurus schistaceus
- Sunda Forktail, Enicurus velatus
- Chestnut-naped Forktail, Enicurus ruficapillus
- Black-backed Forktail, Enicurus immaculatus
- White-crowned Forktail, Enicurus leschenaulti
- Spotted Forktail, Enicurus maculatus
- Blue-fronted Robin, Cinclidium frontale
- Sri Lanka Whistling-Thrush, Myophonus blighi
- Shiny Whistling-Thrush, Myophonus melanurus
- Javan Whistling-Thrush, Myophonus glaucinus
- Brown-winged Whistling-Thrush, Myophonus castaneus
- Bornean Whistling-Thrush, Myophonus borneensis
- Malayan Whistling-Thrush, Myophonus robinsoni
- Malabar Whistling-Thrush, Myophonus horsfieldii
- Taiwan Whistling-Thrush, Myophonus insularis
- Blue Whistling-Thrush, Myophonus caeruleus
- Yellow-rumped Flycatcher, Ficedula zanthopygia
Click for
Ficedula Flycatcher tree - Green-backed Flycatcher, Ficedula elisae
- Narcissus Flycatcher, Ficedula narcissina
- Mugimaki Flycatcher, Ficedula mugimaki
- Slaty-backed Flycatcher, Ficedula sordida
- Slaty-blue Flycatcher, Ficedula tricolor
- Snowy-browed Flycatcher, Ficedula hyperythra
- Pygmy Flycatcher, Ficedula hodgsoni
- Rufous-gorgeted Flycatcher, Ficedula strophiata
- Sapphire Flycatcher, Ficedula sapphira
- Ultramarine Flycatcher, Ficedula superciliaris
- Little Pied Flycatcher, Ficedula westermanni
- Rusty-tailed Flycatcher, Ficedula ruficauda
- Taiga Flycatcher, Ficedula albicilla
- Red-breasted Flycatcher, Ficedula parva
- Kashmir Flycatcher, Ficedula subrubra
- Semicollared Flycatcher, Ficedula semitorquata
- European Pied-Flycatcher, Ficedula hypoleuca
- Collared Flycatcher, Ficedula albicollis
- Atlas Pied-Flycatcher, Ficedula speculigera
- Damar Flycatcher, Ficedula henrici
- Rufous-throated Flycatcher, Ficedula rufigula
- Cinnamon-chested Flycatcher, Ficedula buruensis
- Sumba Flycatcher, Ficedula harterti
- Black-banded Flycatcher, Ficedula timorensis
- Little Slaty Flycatcher, Ficedula basilanica
- Bundok Flycatcher, Ficedula luzoniensis
- Cryptic Flycatcher, Ficedula crypta
- Lompobattang Flycatcher, Ficedula bonthaina
- Black-and-orange Flycatcher, Ficedula nigrorufa
- Tanimbar Flycatcher, Ficedula riedeli
- Rufous-chested Flycatcher, Ficedula dumetoria
- Furtive Flycatcher, Ficedula disposita
- Palawan Flycatcher, Ficedula platenae
- Przevalski's Redstart, Phoenicurus alaschanicus
Click for
Redstart/Rock-Thrush tree - Eversmann's Redstart, Phoenicurus erythronotus
- Blue-fronted Redstart, Phoenicurus frontalis
- Blue-capped Redstart, Phoenicurus coeruleocephala
- White-throated Redstart, Phoenicurus schisticeps
- White-capped Redstart, Phoenicurus leucocephalus
- Plumbeous Water-Redstart, Phoenicurus fuliginosus
- Luzon Water-Redstart, Phoenicurus bicolor
- Black Redstart, Phoenicurus ochruros
- Common Redstart, Phoenicurus phoenicurus
- Moussier's Redstart, Phoenicurus moussieri
- Daurian Redstart, Phoenicurus auroreus
- Gueldenstadt's Redstart, Phoenicurus erythrogastrus
- Hodgson's Redstart, Phoenicurus hodgsoni
- Blue-capped Rock-Thrush, Monticola cinclorhyncha
- Chestnut-bellied Rock-Thrush, Monticola rufiventris
- White-throated Rock-Thrush, Monticola gularis
- Short-toed Rock-Thrush, Monticola brevipes
- Sentinel Rock-Thrush, Monticola explorator
- Littoral Rock-Thrush, Monticola imerina
- Forest Rock-Thrush, Monticola sharpei
- Common Rock-Thrush, Monticola saxatilis
- Blue Rock-Thrush, Monticola solitarius
- Red-bellied Rock-Thrush, Monticola philippensis
- Little Rock-Thrush, Monticola rufocinereus
- White-winged Cliff-Chat, Monticola semirufus
- Miombo Rock-Thrush, Monticola angolensis
- Cape Rock-Thrush, Monticola rupestris
- Jerdon's Bushchat, Saxicola jerdoni
Click for Chat/Wheatear tree - Gray Bushchat, Saxicola ferreus
- White-bellied Bushchat, Saxicola gutturalis
- Whinchat, Saxicola rubetra
- White-browed Bushchat, Saxicola macrorhynchus
- White-throated Bushchat, Saxicola insignis
- Pied Bushchat, Saxicola caprata
- White-tailed Stonechat, Saxicola leucurus
- Stejneger's Stonechat, Saxicola stejnegeri
- Siberian Stonechat, Saxicola maurus
- Canary Islands Stonechat, Saxicola dacotiae
- European Stonechat, Saxicola rubicola
- African Stonechat, Saxicola torquatus
- Madagascan Stonechat, Saxicola sibilla
- Reunion Stonechat, Saxicola tectes
- Buff-streaked Chat, Campicoloides bifasciatus
- Karoo Chat, Emarginata schlegelii
- Sickle-winged Chat, Emarginata sinuata
- Tractrac Chat, Emarginata tractrac
- Moorland Chat, Pinarochroa sordida
- Mocking Cliff-Chat, Thamnolaea cinnamomeiventris
- White-crowned Cliff-Chat, Thamnolaea coronata
- Sooty Chat, Myrmecocichla nigra
- Anteater Chat, Myrmecocichla aethiops
- Congo Moor Chat, Myrmecocichla tholloni
- Ant-eating Chat, Myrmecocichla formicivora
- Mountain Wheatear, Myrmecocichla monticola
- Rueppell's Black Chat, Myrmecocichla melaena
- Arnott's Chat, Myrmecocichla arnotti
- Ruaha Chat, Myrmecocichla collaris
- Northern Wheatear, Oenanthe oenanthe
- Capped Wheatear, Oenanthe pileata
- Red-breasted Wheatear, Oenanthe bottae
- Heuglin's Wheatear, Oenanthe heuglini
- Isabelline Wheatear, Oenanthe isabellina
- Hooded Wheatear, Oenanthe monacha
- Desert Wheatear, Oenanthe deserti
- Black-eared Wheatear, Oenanthe hispanica
- Cyprus Wheatear, Oenanthe cypriaca
- Pied Wheatear, Oenanthe pleschanka
- White-fronted Black Chat, Oenanthe albifrons
- Somali Wheatear, Oenanthe phillipsi
- Red-rumped Wheatear, Oenanthe moesta
- Blackstart, Oenanthe melanura
- Familiar Chat, Oenanthe familiaris
- Brown-tailed Rockchat, Oenanthe scotocerca
- Sombre Rockchat, Oenanthe dubia
- Brown Rockchat, Oenanthe fusca
- Variable Wheatear, Oenanthe picata
- Black Wheatear, Oenanthe leucura
- Abyssinian Wheatear, Oenanthe lugubris
- White-crowned Wheatear, Oenanthe leucopyga
- Hume's Wheatear, Oenanthe albonigra
- Finsch's Wheatear, Oenanthe finschii
- Mourning Wheatear, Oenanthe lugens
- Arabian Wheatear, Oenanthe lugentoides
- “Persian Mourning Wheatear”, Oenanthe persica
- Kurdish Wheatear, Oenanthe xanthoprymna
- Red-tailed Wheatear, Oenanthe chrysopygia
Incertae Sedis: Muscicapidae
It is still quite unclear where the remaining two genera go, so I've put them in a separate Incertae sedis group. They are thought to be in Saxicolinae, but there isn't much real evidence. One or more might belong to Turdidae, or even to a less closely related family. Still, the odds are they go somewhere in Muscicapidae.
- Humblot's Flycatcher, Humblotia flavirostris