Tyrannidae: Tyrant Flycatchers Vigors, 1825
85 genera, 349 species HBW-9
 |
| Click for Tyrannidae species tree |
|---|
The loss of the Rhynchocyclidae and other taxa has reduced the Tyrannidae from their traditional size. Nonetheless, the Tyrannidae remain the largest of the suboscine families. The Tyrannidae include about one quarter of the suboscines.
Although Ohlson et al. (2013) estimate that the Tyrannidae have been a separate lineage since roughly 25-30 million years ago (mid-Oligocene), they found the current diversification of the group dates to the last 20 million years. As usual, there's some dispersion in the age estimates. Harvey et al. (2020) put its origin at 21.8 million years while Oliveros et al. (2019) give about 18.5 mya.
The tyrant flycatchers remain among the most difficult birds to classify and identify, full of cryptic species. The Empidonax flycatchers are an example familiar to North American birders. Some are so cryptic that are often impossible to identity even in the hand, unless they call, such as the Alder (E. alnorum) and Willow Flycathers (E. traillii). With others, such as the Cordilleran (E. occidentalis) and Pacific-slope Flycatchers (E. difficilis), there remains controversy about whether 1 or 2 species are involved. Such problems also occur with tyrant flycatchers in other genera.
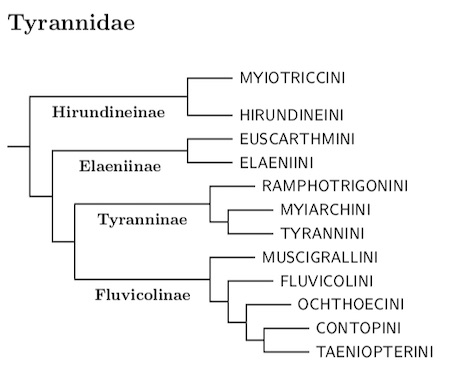 |
| Click for genus-level tree for Tyrannidae |
|---|
The slimmed-down Tyrannidae fall into four main clades, three major and one minor. I rank these clades as subfamilies. The first subamily is Hirundineinae. This is a small subfamily. After some recent splits it still only contains 11 species. Ohlson et al. (2008) considered this group closest to the Tyranninae and Fluvicolinae. In contrast, Fjeldså et al. (2018) and Tello et al. (2009) found them sister to the Elaeniinae. Rheindt et al.'s (2008a) data put them in a more basal position, sister to the combined Elaeniinae, Tyranninae, and Fluvicolinae. Harvey et al. (2020), which I use for the Tyrannidae, have Hirundineinae as the basal clade in Tyrannidae.
The second subfamily contains the Elaenias (Elaeniinae). It split into two major branches about 14 mya. The two branches are the tribes Euscarthmini and Elaeniini. If you prefer more tribes, the split between Zimmerius and the rest of Euscarthmini occurred about 13 mya, and that between the Stigmatura-Inezia and Xanthomyias-Camptostoma groups occurred about 12¼ mya.
The situation in the Elaeniini had been rather complex, with different analyses giving quite different results. Again, Harvey et al. (2020) have resolved many of the issues I had with this group. See Ohlson et al. (2013), Ericson et al. (2006b), Ohlson et al. (2008), Rheindt et al. (2008a), and Tello et al. (2009) for alternative treatments. Within Elaeniini, the next division is at 13 mya, between the Elaenia and Suiriri branches. The latter branch has no natural breakpoints after that.
The remainder of the Tyrannidae breaks into two more clades at 14 mya: Tyranninae and Fluvicolinae. Before July 2021, I left the Tyranninae without any substructure. I now divide them into three tribes. The small Ramphotrigonini branch dates to 13 mya, while the split between Myiarchini (Myiarchus flycatchers and relatives) and Tyrannini (kingbirds and relatives) occurred about 11¼ mya.
The placement of the Short-tailed Field Tyrant, Muscigralla brevicauda, was previously somewhat soft and I put it in its own subfamily because I wasn't sure where it belonged. In Harvey et al. (2020) the tyrant clearly groups with the Fluvicolinae and I regard it as forming the first of the Fluvicoline tribes: Muscigrallini. This branch dates to about 13.7 mya.
The other four tribes I recognize in Fluvicolinae date from 12.5-12.0 mya. Of them, Ochthoecini is the oldest, followed by Fluvicolini. The other two, Contopini and Taeniopterini (sometimes called Xolmiini or Xolmini) are the youngest at 12 mya. I considered adding one or two more, but in the end, didn't think it was informative enough to justify the additional compliction.
Hirundineinae Tello et al., 2009
Notes — Hirundineinae
The Hirundineinae have grown to 11 species due to splits based on Harvey et al. (2020) and the HBW Checklist (del Hoyo and Collar, 2016). These splits involve three species.
Ornate Flycatcher Split: Based on Harvey et al. (2020), who appear to have sampled phoenicurus and ornatus, and the comments in the HBW Checklist, the Ornate Flycatcher, Myiotriccus ornatus, is split into
- Eastern Ornate-Flycatcher, Myiotriccus phoenicurus + aureiventris
- Western Ornate-Flycatcher, Myiotriccus ornatus + stellatus
Former Myiophobus: The three former Myiophobus included in this subfamily are not closely related to the rest of Myiophobus. Ohlsson et al. (2009) created a new genus name for them, Nephelomyias.
Cliff Flycatcher Split: Based on Harvey et al. (2020) showing about 2 million years separation and the comments in the HBW Checklist, Cliff Flycatcher, Hirundinea ferruginea is split into
- Cliff Flycatcher, Hirundinea ferruginea inc. sclateri
- Swallow Flycatcher, Hirundinea bellicosa inc. pallidior
Cinnamon Flycatcher Split: Harvey et al. (2020) found distances of 5-7 million years between three subspecies of Cinnamon Flycatcher (Pyrrhomyias cinnamomeus), including the two subspecies in Sucre, Venezuela. When combined with the information in the HBW Checklist, Cinnamon Flycatcher has been split into
- Santa Marta Cinnamon-Flycatcher, Pyrrhomyias assimilis
- Paria Cinnamon-Flycatcher, Pyrrhomyias pariae
- Venezuelan Cinnamon-Flycatcher, Pyrrhomyias vieillotioides + spadix
- Andean Cinnamon-Flycatcher, Pyrrhomyias cinnamomeus + pyrrhopterus
Hirundineinae Tello et al., 2009
Myiotriccini: Ornate and Handsome Flycatchers Informal
- Eastern Ornate-Flycatcher, Myiotriccus phoenicurus
- Western Ornate-Flycatcher, Myiotriccus ornatus
- Handsome Flycatcher, Nephelomyias pulcher
- Orange-banded Flycatcher, Nephelomyias lintoni
- Ochraceous-breasted Flycatcher, Nephelomyias ochraceiventris
Hirundineini:Cliff and Cinnamon Flycatchers Tello et al., 2009
- Cliff Flycatcher, Hirundinea ferruginea
- Swallow Flycatcher, Hirundinea bellicosa
- Andean Cinnamon-Flycatcher, Pyrrhomyias cinnamomeus
- Santa Marta Cinnamon-Flycatcher, Pyrrhomyias assimilis
- Venezuelan Cinnamon-Flycatcher, Pyrrhomyias vieillotioides
- Paria Cinnamon-Flycatcher, Pyrrhomyias pariae
Elaeniinae: Elaenine Flycatchers Cabanis & Heine, 1859-60
There are two tribes: Euscarthmini and Elaeniini.
Notes — Euscarthmini
Besides Harvey et al. (2020), the arrangement of species within Zimmerius is also based on Rheindt et al. (2008c, d, 2013). Based on Rheindt et al. (2013), Mistletoe Tyrannulet, Zimmerius parvus, Specious Tyrannulet, Zimmerius improbus (inc. tamae), and Venezuelan Tyrannulet, Zimmerius petersi, are split from Paltry Tyrannulet, Zimmerius vilissimus.
Coopmans's Tyrannulet, Zimmerius minimus, comprising minimus and cumanensis has somewhat speculatively been split from Golden-faced Tyrannulet, Zimmerius chrysops, based on comments in Rheindt et al., (2013). I continue include the Loja Tyrannulet, Zimmerius flavidifrons. but Rheindt et al., (2014) describe a mosiac population that is linked to both the Golden-faced and Peruvian Tyrannulets. It includes flavidifrons and an undescribed Peruvian population and may end up being lumped in with the Peruvian Tyrannulet. Thus the old chrysops with 5 subspecies has been divided among Coopmans's, Choco, Golden-faced, and Loja Tyrannulets. Finally, Chico's Tyrannulet, Zimmerius chicomendesi, newly described by Whitney et al., (2013c), has been added to the list.
Gray-capped Tyrannulet: The Gray-capped Tyrannulet, formerly Phyllomyias griseocapilla, has been transferred to Zimmerius. I dithered some on this.
Given that its common ancestor with the Zimmerius tyrannulets was about 8.7 mya, my first reaction was to list its genus as "Phyllomyias". I had only seen one once, back in 2010, and couldn't recall what it looked like. The illustration in the HBW Checklist didn't help, so I checked photos via Birds of the World. They didn't look much like the illustration, and showed a Zimmerius-like wing pattern. That impressed me enough to put it in Zimmerius instead.
Here's an excerpt from Traylor (1977) where he defines Zimmerius.
Diagnosis. Superficially most like Tyranniscus Cabanis and Heine, 1859, but differs as follows: has a unique wing pattern in which the median and greater coverts, the secondaries and four or five subexternal primaries are edged with yellow, while the outermost and four or five internal primaries are blackish, producing a black wedge when the wing is spread
Traylor named the genus after John Todd Zimmer partly because of his recognition of the unique wing pattern in the birds Traylor named Zimmerius.
Wagtail-Tyrants Split: Based on Harvey et al. (2020) and the HBW Checklist, there are two splits in the wagtail-tyrants (Stigmatura):
- The Lesser Wagtail-Tyrant, Stigmatura napensis, is split into
- Lesser Wagtail-Tyrant, Stigmatura napensis
- Bahia Wagtail-Tyrant, Stigmatura bahiae.
- Greater Wagtail-Tyrant, Stigmatura budytoides, is split into
- Greater Wagtail-Tyrant, Stigmatura budytoides
- Caatinga Wagtail-Tyrant, Stigmatura gracilis.
Tawny-crowned Pygmy-Tyrant: Following SACC #898 and Franz et al. (2020), the Tawny-crowned Pygmy-Tyrant, Euscarthmus meloryphus, has been split into
- Tawny-fronted Pygmy-Tyrant, Euscarthmus fulviceps
- Tawny-crowned Pygmy-Tyrant, Euscarthmus meloryphus.
Sixteen species of Phyllomyias tyrannulets were earlier separated from the main Phyllomyias group (part of Elaeniini) and placed variously in the revived genera Tyranniscus and Xanthomyias. I've also included five tyrannulets from Mercocerculus in Xanthomyias. This reduces Mercocerculus itself to a single species which belongs in Elaeniini.
Beardless-Tyrannulets: Based on Fitzpatrick (2004a), Rheindt et al. (2008d), Harvey et al. (2020), and the HBW Checklist, the Southern Beardless-Tyrannulet, Camptostoma obsoletum, has been split into five species:
- Western Beardless-Tyrannulet, Camptostoma sclateri, including maranonicum and griseum
- Central American Beardless-Tyrannulet, Camptostoma flaviventre, including orphnum and majus
- Colombian Beardless-Tyrannulet, Camptostoma pusillum, including napaeum and perhaps caucae
- Olive Beardless-Tyrannulet, Camptostoma olivaceum, monotypic
- Southern Beardless-Tyrannulet, Camptostoma obsoletum, including bolivianum and cinerascens
Ever since I saw them in different parts of South America, I've wondered whether the Southern Beardless-Tyrannulets were all the same species. Looks unlikely now! The oldest branch is about 6 million years old. If you examine the tree, you'll see that the Northern Beardless-Tyrannulet, Camptostoma imberbe is embedded in this group. I've used names from the HBW Checklist. Of them, the name Colombian Beardless-Tyrannulet is somewhat misleading as its range extends far eastward, including the Guianas and adjacent regions of Brazil north of the Amazon. Beardless-Tyrannulets in Maranhão east and south (cinerascens) are in the Southern group. Concerning caucae, see the speculation by Rheindt et al. (2008d). This group could use more study.
Notes — Elaeniini
Chapada Flycatcher: The Chapada Flycatcher, Suiriri affinis, is unrelated to the other Suiriri taxa and has been removed to Fluvicolini.
Nesotriccus: Zucker et al. (2016) found that Nesotriccus is embedded in Phaeomyias and that both are very closely related to Phyllomyias. Accordingly, I have merged Nesotriccus (Cocos Flycatcher) and Phaeomyias into Phyllomyias. It would not be unreasonable to go a step further and merge the monotypic Capsiempis with Phyllomyias. However, both names have equal priority and apparently need a first reviser action to decide between them. Since there's not a strong need to merge them, it can wait.
Mouse-colored Tyrannulet: Also based on Zucker et al. (2016), the Mouse-colored Tyrannulet, formerly Phaeomyias murina, has been split into Amazonian Mouse-colored Tyrannulet, Phyllomyias murinus, including wagae, and Northern Mouse-colored Tyrannulet, Phyllomyias incomtus, including eremonomus.
Straneck's Tyrannulet: Straneck's Tyrannulet / Gray-crowned Tyrannulet, Serpophaga griseicapilla, has had a somewhat checkered history. It was previously referred to as Serpophaga griseiceps, but the type of griseiceps was actually a juvenile of S. munda (Herzog and Mazur Barnett, 2004). Straneck (2007) gave it the name S. griseiceps. It has also been referred to as Gray-crowned Tyrannulet, and Monte Tyrannulet.
More Elaenias: The ordering within Elaenia itself is now based on Harvey et al. (2020), which has some minor differences from Tang et al. (2018), which I used previously. Rheindt et al. (2008b, 2009a) indicated that the Tepui Elaenia should be split from the Sierran Elaenia; the Greater Antillean Elaenia split into Sclater's Elaenia, Elaenia fallax, of Jamaica and the Hispaniolan Elaenia, Elaenia cherriei. Rheindt et al. (2008b, 2009a) also found evidence for splitting the Chilean Elaenia from the Sierran Elaenia, but the additional data studied by Tang et al. (2018) suggests they cannot be separated.
Although E. fallax used to be called Jamaican Elaenia, that name is now used for Myiopagis cotta, formerly known as the Cotta Flycatcher, after Cotta Wood, which was near Content, Jamaica.
Not all subspecies of the White-crested Elaenia group were analyzed, but the following is a plausible arrangement of the relevant subspecies: Tepui Elaenia, Elaenia olivina, includes davidwillardi and White-crested Elaenia, Elaenia albiceps, includes griseigularis, diversa, urubambae, and modesta. Although it was not part of Tang et al.'s analysis, modesta is distinctive. Harvey et al. (2020) did include one in their analysis, and I have used that to support a split. Finally, Tang et al. found that the subspecies sorida should be split from the Highland Elaenia E. obscura. This subspecies, has now been split as Small-headed Elaenia. Coopmans's Elaenia, Elaenia brachyptera, has been split from Lesser Elaenia, Elaenia chiriquensis, based on Rheindt et al. (2015). Finally, the Myiopagis Elaenias are organized as in Rheindt et al. (2008b, 2009a)
Gray Elaenia: Based on Harvey et al. (2020) and the HBW Checklist, the Gray Elaenia, Myiopagis caniceps has been split into three species:
- Choco Gray Elaenia, Myiopagis parambae, includes absita
- Amazonian Gray Elaenia, Myiopagis cinerea
- Atlantic Gray Elaenia, Myiopagis caniceps
Greenish Elaenia: Based on Harvey et al. (2020) and the HBW Checklist, the Greenish Elaenia, Myiopagis viridicata has been split into two species:
- West Mexican Elaenia, Myiopagis minima, including jaliscensis
- Greenish Elaenia, Myiopagis viridicata, all other subspecies
Small-headed Elaenia: Based on Harvey et al. (2020), SACC #806, and SACC #813, the Highland Elaenia, Elaenia obscura, has been split into
- Small-headed Elaenia, Elaenia sordida
- Highland Elaenia, Elaenia obscura
White-crested Elaenia: Based on Harvey et al. (2020) and the HBW Checklist, the White-crested Elaenia, Elaenia albiceps, has been split into three species:
- White-crested Elaenia, Elaenia albiceps
- Peruvian Elaenia, Elaenia modesta
- Chilean Elaenia, Elaenia chilensis
I've gone back and forth on the Chilean Elaenia. Harvey et al. made clear that is it is genetically quite different from albiceps. I don't have any genetic information on the Peruvian Elaenia, but it seems like a good bet.
Ridgwayornis: I've separated two Serpophaga in genus Ridgwayornis (Bertoni 1925), type nigricans.
- Torrent Tyrannulet, Serpophaga cinerea
- Sooty Tyrannulet, Serpophaga nigricans
The two Tachuri (Polystictus): The Bearded Tachuri, Polystictus pectoralis, and Gray-backed Tachuri, Polystictus superciliaris, are not sister species. This forces the removal of the Gray-backed Tachuri to Serpophaga. Given the tree in Harvey et al. (2020) and the similarity of the two Tachuris, it makes sense to transfer both Polystictus to Serpophaga.
Elaeniinae: Elaenine Flycatchers Cabanis & Heine, 1859-60
Euscarthmini: Tyrannulets von Ihering, 1904
- Gray-capped Tyrannulet, Zimmerius griseocapilla
- Red-billed Tyrannulet, Zimmerius cinereicapilla
- Mishana Tyrannulet, Zimmerius villarejoi
- Chico's Tyrannulet, Zimmerius chicomendesi
- Spectacled Tyrannulet, Zimmerius improbus
- Venezuelan Tyrannulet, Zimmerius petersi
- Bolivian Tyrannulet, Zimmerius bolivianus
- Slender-footed Tyrannulet, Zimmerius gracilipes
- Guianan Tyrannulet, Zimmerius acer
- Paltry Tyrannulet / Guatemalan Tyrannulet, Zimmerius vilissimus
- Mistletoe Tyrannulet, Zimmerius parvus
- Coopmans's Tyrannulet, Zimmerius minimus
- Choco Tyrannulet, Zimmerius albigularis
- Golden-faced Tyrannulet, Zimmerius chrysops
- Loja Tyrannulet, Zimmerius flavidifrons
- Peruvian Tyrannulet, Zimmerius viridiflavus
- Lesser Wagtail-Tyrant, Stigmatura napensis
- Caatinga Wagtail-Tyrant, Stigmatura gracilis
- Greater Wagtail-Tyrant, Stigmatura budytoides
- Bahia Wagtail-Tyrant, Stigmatura bahiae
- Tawny-crowned Pygmy-Tyrant, Euscarthmus meloryphus
- Tawny-fronted Pygmy-Tyrant, Euscarthmus fulviceps
- Rufous-sided Pygmy-Tyrant, Euscarthmus rufomarginatus
- Pale-tipped Tyrannulet / Pale-tipped Inezia, Inezia caudata
- Amazonian Tyrannulet / Amazonian Inezia, Inezia subflava
- Slender-billed Tyrannulet / Slender-billed Inezia, Inezia tenuirostris
- Plain Tyrannulet / Plain Inezia, Inezia inornata
- Sulphur-bellied Tyrannulet, Xanthomyias minor
- Rufous-winged Tyrannulet, Xanthomyias calopterus
- White-banded Tyrannulet, Xanthomyias stictopterus
- Buff-banded Tyrannulet, Xanthomyias hellmayri
- White-tailed Tyrannulet, Xanthomyias poecilocercus
- Plumbeous-crowned Tyrannulet, Xanthomyias plumbeiceps
- Greenish Tyrannulet, Xanthomyias virescens
- Sclater's Tyrannulet, Xanthomyias sclateri
- Urich's Tyrannulet, Xanthomyias urichi
- Reiser's Tyrannulet, Xanthomyias reiseri
- White-fronted Tyrannulet, Tyranniscus zeledoni
- Rough-legged Tyrannulet, Tyranniscus burmeisteri
- Ashy-headed Tyrannulet, Tyranniscus cinereiceps
- Black-capped Tyrannulet, Tyranniscus nigrocapillus
- Tawny-rumped Tyrannulet, Tyranniscus uropygialis
- White-lored Tyrannulet, Ornithion inerme
- Yellow-bellied Tyrannulet, Ornithion semiflavum
- Brown-capped Tyrannulet, Ornithion brunneicapillus
- Western Beardless-Tyrannulet, Camptostoma sclateri
- Northern Beardless-Tyrannulet, Camptostoma imberbe
- Central American Beardless-Tyrannulet, Camptostoma flaviventre
- Colombian Beardless-Tyrannulet, Camptostoma pusillum
- Olive Beardless-Tyrannulet, Camptostoma olivaceum
- Southern Beardless-Tyrannulet, Camptostoma obsoletum
Elaeniini: Elaenias, Tit-Tyrants Cabanis & Heine, 1859-60
- Yellow-crowned Tyrannulet, Tyrannulus elatus
- Choco Gray Elaenia, Myiopagis parambae
- Foothill Elaenia, Myiopagis olallai
- Amazonian Gray Elaenia, Myiopagis cinerea
- Atlantic Gray Elaenia, Myiopagis caniceps
- Forest Elaenia, Myiopagis gaimardii
- Pacific Elaenia, Myiopagis subplacens
- Yellow-crowned Elaenia, Myiopagis flavivertex
- West Mexican Elaenia, Myiopagis minima
- Greenish Elaenia, Myiopagis viridicata
- Jamaican Elaenia, Myiopagis cotta
- Plain-crested Elaenia, Elaenia cristata
- Rufous-crowned Elaenia, Elaenia ruficeps
- Slaty Elaenia, Elaenia strepera
- Mottle-backed Elaenia, Elaenia gigas
- Small-headed Elaenia, Elaenia sordida
- Great Elaenia, Elaenia dayi
- Yellow-bellied Elaenia, Elaenia flavogaster
- Highland Elaenia, Elaenia obscura
- Small-billed Elaenia, Elaenia parvirostris
- Coopmans's Elaenia, Elaenia brachyptera
- Brownish Elaenia, Elaenia pelzelni
- Large Elaenia, Elaenia spectabilis
- Noronha Elaenia, Elaenia ridleyana
- Lesser Elaenia, Elaenia chiriquensis
- Olivaceous Elaenia, Elaenia mesoleuca
- Sierran Elaenia, Elaenia pallatangae
- White-crested Elaenia, Elaenia albiceps
- Peruvian Elaenia, Elaenia modesta
- Chilean Elaenia, Elaenia chilensis
- Tepui Elaenia, Elaenia olivina
- Mountain Elaenia, Elaenia frantzii
- Caribbean Elaenia, Elaenia martinica
- Sclater's Elaenia, Elaenia fallax
- Hispaniolan Elaenia, Elaenia cherriei
- Suiriri Flycatcher, Suiriri suiriri
- Gray-and-white Tyrannulet, Pseudelaenia leucospodia
- Yellow Tyrannulet, Capsiempis flaveola
- Sooty-headed Tyrannulet, Phyllomyias griseiceps
- Yungas Tyrannulet, Phyllomyias weedeni
- Planalto Tyrannulet, Phyllomyias fasciatus
- Tumbesian Tyrannulet, Phyllomyias tumbezanus
- Amazonian Mouse-colored Tyrannulet, Phyllomyias murinus
- Cocos Flycatcher, Phyllomyias ridgwayi
- Northern Mouse-colored Tyrannulet, Phyllomyias incomtus
- White-throated Tyrannulet, Mecocerculus leucophrys
- Sharp-tailed Tyrant / Sharp-tailed Grass-Tyrant, Culicivora caudacuta
- Torrent Tyrannulet, Ridgwayornis cinereus
- Sooty Tyrannulet, Ridgwayornis nigricans
- Crested Doradito, Pseudocolopteryx sclateri
- Subtropical Doradito, Pseudocolopteryx acutipennis
- Dinelli's Doradito, Pseudocolopteryx dinelliana
- Ticking Doradito, Pseudocolopteryx citreola
- Warbling Doradito, Pseudocolopteryx flaviventris
- Bearded Tachuri, Serpophaga pectoralis
- Gray-backed Tachuri, Serpophaga superciliaris
- Straneck's Tyrannulet / Gray-crowned Tyrannulet, Serpophaga griseicapilla
- River Tyrannulet, Serpophaga hypoleuca
- White-bellied Tyrannulet, Serpophaga munda
- White-crested Tyrannulet, Serpophaga subcristata
- Agile Tit-Tyrant, Uromyias agilis
- Unstreaked Tit-Tyrant, Uromyias agraphia
- Black-crested Tit-Tyrant, Anairetes nigrocristatus
- Pied-crested Tit-Tyrant, Anairetes reguloides
- Ash-breasted Tit-Tyrant, Anairetes alpinus
- Yellow-billed Tit-Tyrant, Anairetes flavirostris
- Tufted Tit-Tyrant, Anairetes parulus
- Juan Fernandez Tit-Tyrant, Anairetes fernandezianus
Tyranninae: Tyrant Flycatchers Vigors, 1825
The subfamily Tyranninae is now divided into three tribes: Ramphotrigonini, Myiarchini, and Tyrannini.
Notes — Myiarchini
Pale-bellied Mourner: Based on Harvey et al. (2020), the Pale-bellied Mourner, Rhytipterna immunda has been transferred to the genus Casiornis.
Grayish Mourner: Based on Harvey et al. (2020), the Grayish Mourner, Rhytipterna simplex, is split into:
- Amazonian Grayish Mourner, Rhytipterna frederici
- Atlantic Grayish Mourner, Rhytipterna simplex
The reason for the split is that the Atlantic Grayish Mourner's closest relative is the Rufous Mourner, Rhytipterna holerythra.
Rufous Flycatcher: The Rufous Flycatcher, Myiarchus semirufus, has been transferred to the monotypic genus Muscifur (Bangs and Penard, 1921). While the other Myiarchus have a common ancestor a bit over 4 million years ago, their common ancestor with the Rufous Flycatcher is close to 8 mya. Moreover, with its dark cap and rufous coloration, the Rufous Flycatcher looks a little bit like a Casiornis and a little bit like a Sirystes. I'm happier having it in its own genus.
Myiarchus: The arrangement of Myiarchus is based on Harvey et al. (2020), with some help from Joseph et al. (2004) and Sari and Parker (2012). They later two note some issues within Myiarchus. They may represent incompelete lineage sorting, species boundaries that need to be redrawn, or even cryptic species. One such involves La Sagra's and Stolid Flycatchers, which seem to be mutually paraphyletic (Joseph et al. 2004). Other such problems exist for ferox, phaeocephalus, swainsoni, and tuberculifer.
Nutting's Flycatcher: Based on Harvey et al. (2020) and the HBW Checklist, Nutting's Flycatcher, Myiarchus nuttingi, has been split into:
- Nutting's Flycatcher, Myiarchus nuttingi, including inquietus
- Ridgway's Flycatcher, Myiarchus flavidior
Brown-crested Flycatcher: Based on Harvey et al. (2020), Joseph et al. (2004), and Sari and Parker (2012), I've split the Brown-crested Flycatcher, Myiarchus tyrannulus, into two species:
- Northern Brown-crested Flycatcher, Myiarchus cooperi, including magister, cozumelae, insularum, and brachyurus.
- Southern Brown-crested Flycatcher, Myiarchus tyrannulus, including bahiae
The HBW Checklist mentions three subspecies groups within cooperi. Combining Joseph et al. and Sari and Parker suggests that the northern group forms a clade (neither paper includes all the pieces). The splits made are due to the relation with the Granada and Galapagos Flycatchers per Harvey et al. The genetic distance is in the ambiguous range for splits, so I have not adopted any others.
Sirystes: Based on Donegan (2013), Sirystes, Sirystes sibilator, has been split into:
- Choco Sirystes, Sirystes albogriseus
- Todd's Sirystes, Sirystes subcanescens
- White-rumped Sirystes, Sirystes albocinereus
- Sibilant Sirystes, Sirystes sibilator
Notes — Tyrannini
Lesser Kiskadee: I've put the Lesser Kiskadee in Philohydor as it appears to not be on the same branch as Pitangus. They are different branches in Harvey et al. (2020). They are sister species in Tello et al. (2009), but the support is not great and Chaves et al. (2008) has them separated.
Milvulus: I've also separated the Fork-tailed Flycatcher in Milvulus based on Tello et al. (2009). Once it is out of Tyrannus it reclaims its old name and becomes Milvulus tyrannus, not Milvulus savana.
Loggerhead Kingbird: Based on Garrido et al. (2009), the Loggerhead Kingbird has been split into three species:
- Western Loggerhead Kingbird, Tyrannus caudifasciatus
- Hispaniolan Kingbird, Tyrannus gabbii
- Puerto Rican Kingbird, Tyrannus taylori
The three are easily distinguished by their calls, or with a little care, by their plumage.
Streaked Flycatcher: Based on Harvey et al. (2020) and the HBW Checklist, and presuming their maculatus sample from Loreto is a wintering solitarius (otherwise, even more revision is needed), the Streaked Flycatcher, Myiodynastes maculatus, is split into two species:
- Northern Streaked-Flycatcher, Myiodynastes maculatus
- Southern Streaked-Flycatcher, Myiodynastes solitarius (monotypic)
PS. Still waiting for the Island Streaked-Flycatcher to be described…
Tyranninae: Tyrant Flycatchers Vigors, 1825
Ramphotrigonini: Flatbills Informal
- Large-headed Flatbill, Ramphotrigon megacephalum
- Rufous-tailed Flatbill, Ramphotrigon ruficauda
- Flammulated Flycatcher, Ramphotrigon flammulatum
- Dusky-tailed Flatbill, Ramphotrigon fuscicauda
Myiarchini: Myiarchus Flycatchers von Berlepsch, 1907
- Choco Sirystes, Sirystes albogriseus
- Todd's Sirystes, Sirystes subcanescens
- White-rumped Sirystes, Sirystes albocinereus
- Sibilant Sirystes, Sirystes sibilator
- Amazonian Grayish Mourner, Rhytipterna frederici
- Rufous Mourner, Rhytipterna holerythra
- Atlantic Grayish Mourner, Rhytipterna simplex
- Pale-bellied Mourner, Casiornis immunda
- Rufous Casiornis, Casiornis rufus
- Ash-throated Casiornis, Casiornis fuscus
- Rufous Flycatcher, Muscifur semirufus
- Swainson's Flycatcher, Myiarchus swainsoni
- Dusky-capped Flycatcher, Myiarchus tuberculifer
- Sad Flycatcher, Myiarchus barbirostris
- Venezuelan Flycatcher, Myiarchus venezuelensis
- Pale-edged Flycatcher, Myiarchus cephalotes
- Apical Flycatcher, Myiarchus apicalis
- Sooty-crowned Flycatcher, Myiarchus phaeocephalus
- Panama Flycatcher / Panamanian Flycatcher, Myiarchus panamensis
- Short-crested Flycatcher, Myiarchus ferox
- Rufous-tailed Flycatcher, Myiarchus validus
- Yucatan Flycatcher, Myiarchus yucatanensis
- Ash-throated Flycatcher, Myiarchus cinerascens
- Nutting's Flycatcher, Myiarchus nuttingi
- Ridgway's Flycatcher, Myiarchus flavidior
- Northern Brown-crested Flycatcher, Myiarchus cooperi
- Galapagos Flycatcher, Myiarchus magnirostris
- Southern Brown-crested Flycatcher, Myiarchus tyrannulus
- Grenada Flycatcher, Myiarchus nugator
- Great Crested Flycatcher, Myiarchus crinitus
- La Sagra's Flycatcher, Myiarchus sagrae
- Stolid Flycatcher, Myiarchus stolidus
- Puerto Rican Flycatcher, Myiarchus antillarum
- Lesser Antillean Flycatcher, Myiarchus oberi
Tyrannini: Tyrant Flycatchers Vigors, 1825
- Piratic Flycatcher, Legatus leucophaius
- Bright-rumped Attila, Attila spadiceus
- Cinnamon Attila, Attila cinnamomeus
- Rufous-tailed Attila, Attila phoenicurus
- Ochraceous Attila, Attila torridus
- Citron-bellied Attila, Attila citriniventris
- Dull-capped Attila / White-eyed Attila, Attila bolivianus
- Gray-hooded Attila, Attila rufus
- Cattle Tyrant, Machetornis rixosa
- Lesser Kiskadee, Philohydor lictor
- Great Kiskadee, Pitangus sulphuratus
- Rusty-margined Flycatcher, Myiozetetes cayanensis
- Social Flycatcher, Myiozetetes similis
- Gray-capped Flycatcher, Myiozetetes granadensis
- Dusky-chested Flycatcher, Myiozetetes luteiventris
- Baird's Flycatcher, Myiodynastes bairdii
- Golden-bellied Flycatcher, Myiodynastes hemichrysus
- Golden-crowned Flycatcher, Myiodynastes chrysocephalus
- Sulphur-bellied Flycatcher, Myiodynastes luteiventris
- Northern Streaked-Flycatcher, Myiodynastes maculatus
- Southern Streaked-Flycatcher, Myiodynastes solitarius
- Boat-billed Flycatcher, Megarynchus pitangua
- Sulphury Flycatcher, Tyrannopsis sulphurea
- White-ringed Flycatcher, Conopias albovittatus
- Yellow-throated Flycatcher, Conopias parvus
- Lemon-browed Flycatcher, Conopias cinchoneti
- Three-striped Flycatcher, Conopias trivirgatus
- Variegated Flycatcher, Empidonomus varius
- Crowned Slaty Flycatcher, Empidonomus aurantioatrocristatus
- Fork-tailed Flycatcher, Milvulus tyrannus
- White-throated Kingbird, Tyrannus albogularis
- Tropical Kingbird, Tyrannus melancholicus
- Gray Kingbird, Tyrannus dominicensis
- Giant Kingbird, Tyrannus cubensis
- Snowy-throated Kingbird, Tyrannus niveigularis
- Western Loggerhead Kingbird, Tyrannus caudifasciatus
- Hispaniolan Kingbird, Tyrannus gabbii
- Puerto Rican Kingbird, Tyrannus taylori
- Thick-billed Kingbird, Tyrannus crassirostris
- Couch's Kingbird, Tyrannus couchii
- Eastern Kingbird, Tyrannus tyrannus
- Cassin's Kingbird, Tyrannus vociferans
- Western Kingbird, Tyrannus verticalis
- Scissor-tailed Flycatcher, Tyrannus forficatus
Fluvicolinae: Fluvicoline Flycatchers Swainson, 1832-33
The subfamily Fluvicolinae is divided into five tribes, the monotypic Muscigrallini, Ochthoecini, Fluvicolini, Contopini, and Taeniopterini.
Notes — Ochthoecini
Previous editions of the TiF list had separated Silvicultrix from Ochthoeca. It remains a good idea! The genus Tumbezia was merged into Ochthoeca, with Ohlson et al. (2013) pushing me over the line on this.
"Myiophobus": The temporary genus name "Myiophobus" has been replaced by Scotomyias (Ohlson et al., 2020).
Flavescent Flycatcher: Based on Harvey et al. (2020) and the HBW CHecklist, the Flavescent Flycatcher, Scotomyias flavicans, is split into:
- Flavescent Flycatcher, Scotomyias flavicans
- Haughty Flycatcher, Scotomyias superciliosus (monotypic)
Yes, I made up the name Haughty Flycatcher. It seemed too good to resist.
Chat-Tyrants: Based on Harvey et al. (2020) and the HBW CHecklist, the Crowned Chat-Tyrant, Silvicultrix frontalis, has been split into
- Crowned Chat-Tyrant, Silvicultrix frontalis, including albidiadema
- Kalinowski's Chat-Tyrant, Silvicultrix spodionota, including boliviana
Previously, I split the Slaty-backed Chat-Tyrant, Ochthoeca cinnamomeiventris into:
- Maroon-belted Chat-Tyrant, Ochthoeca thoracica
- Slaty-backed Chat-Tyrant, Ochthoeca cinnamomeiventris
- Blackish Chat-Tyrant, Ochthoeca nigrita
This was based on a combination of Fjeldså et al. (2018), Ridgely and Greenfield (2001), Hilty (2003), and Garcia-Moreno et al. (1998). This is also supported by Harvey et al. (2020), where the three cinnamomeiventris in their tree are the three species.
Notes — Fluvicolini
Following Fjeldså et al. (2018), I've included Colonia here, as well as the Black-and-white Monjita, now called Heteroxolmis dominicanus, previously in genus Xolmis. There is one other addition. The Chapada Flycatcher, formerly Suiriri affinis (not S. islerorum, see Kirwan et al., 2014a) is not related to Suiriri. Rather, Lopes et al. (2018) found it is sister to Sublegatus. Lopes et al. established a new genus for it, Guyramemua, so the Chapada Flycatcher becomes Guyramemua affine and moves from Elaeniini to Fluvicolini.
Vermilion Flycatcher: Based on Carmi et al. (2016), the Vermilion Flycatcher, formerly Pyrocephalus rubinus, has been split into four species:
- Vermilion Flycatcher, now called Pyrocephalus obscurus
- Scarlet Flycatcher, Pyrocephalus rubinus
- Darwin's Flycatcher, Pyrocephalus nanus
- the extinct San Cristobal Flycatcher, Pyrocephalus dubius
Notes — Contopini
Tufted Flycatcher: Based on Harvey et al., (2020) and the HBW Checklist, the Tufted Flycatcher, Mitrephanes phaeocercus, has been split into:
- Mexican Tufted-Flycatcher, Mitrephanes phaeocercus, including tenuirostris
- Costa Rican Tufted-Flycatcher, Mitrephanes aurantiiventris
- Choco Tufted-Flycatcher, Mitrephanes berlepschi including eminulus
Two of the three species were included in Harvey et al. (2020).
Pewees: Based on Harvey et al. (2020) and the HBW CHecklist, the Tropical Pewee, Contopus cinereus, has been split into
- Northern Tropical Pewee, Contopus bogotensis
- Tumbes Pewee, Contopus punensis
- Southern Tropical Pewee, Contopus cinereus.
Notes — Taeniopterini
The more complete taxon sampling by Fjeldså et al. (2018) has forced a split-up of the genus Xolmis. Besides sending the Black-and-white Monjita to the Fluvicolini (Heteroxolmis), The Fire-eyed Diucon, now Pyrope pyrope, moves to Pyrope. I've also moved the Black-crowned Monjita, Neoxolmis coronatus, Rusty-backed Monjita, Neoxolmis rubetra, and Salinas Monjita, Neoxolmis salinarum, to Neoxolmis from Xolmis. Finally, the Gray Monjita, now Nengetus cinereus, has been placed in Nengetus (Swainson 1827). It had historically been in the genus Taenioptera (Bonaparte 1825), of which it is the type. However, although Bonaparte alluded to Taenioptera as a subgenus in 1825, he did not actually establish the name until 1830. Thus Nengetus has priority and Taenioptera is a junior synonym.
Taeniopterini replaces Xolmiini/Xolmini: Besides naming the genus Taenioptera, Bonaparte also created a valid subfamily name based on Taenioptera Taeniopterinae in 1838, which we use in the form Taeniopterini for this tribe. The type species for Bonaparte's Taeniopterinae is Nengetus cinereus = Tyrannus cinereus Vieillot 1816 = Muscicapa taenioptera Bonaparte 1825. This means Taeniopterini has priority over the recent Xolmini (Xolmiini).
Little Ground-Tyrant:
The Little Ground-Tyrant, Muscisaxicola fluviatilis, has been transferred
to genus Syrtidicola, (Chesser et al., 2020b).
[Fluvicolinae, Tyrannida II, 3.07]
Black-Tyrants: Since Eumyiobius would be embedded in Knipolegus, it has been merged into Knipolegus. Hosner and Moyle's (2012) results provide support for treating Caatinga Black-Tyrant, Knipolegus franciscanus, as a separate species from White-winged Black-Tyrant, Knipolegus aterrimus, and also Plumbeous Tyrant, Knipolegus cabanisi, separate from Jelski's Black-Tyrant, Knipolegus signatus.
Based on Harvey et al. (2020) and the HBW CHecklist, the White-winged Black-Tyrant, Knipolegus aterrimus, has been split into:
- White-rumped Black-Tyrant, Knipolegus heterogyna (monotypic)
- White-winged Black-Tyrant, Knipolegus aterrimus
Harvey et al. (2020) found that they are not even sister species.
Fluvicolinae: Fluvicoline Flycatchers Swainson, 1832-33
Muscigrallini: Field Tyrant Ohlson et al., 2013
- Short-tailed Field Tyrant, Muscigralla brevicauda
Ochthoecini: Chat-Tyrants Ohlson et al., 2020
- Bran-colored Flycatcher, Myiophobus fasciatus
- Olive-chested Flycatcher, Myiophobus cryptoxanthus
- Orange-crested Flycatcher, Scotomyias phoenicomitra
- Roraiman Flycatcher, Scotomyias roraimae
- Flavescent Flycatcher, Scotomyias flavicans
- Haughty Flycatcher, Scotomyias superciliosus
- Unadorned Flycatcher, Scotomyias inornatus
- Crowned Chat-Tyrant, Silvicultrix frontalis
- Kalinowski's Chat-Tyrant, Silvicultrix spodionota
- Yellow-bellied Chat-Tyrant, Silvicultrix diadema
- Jelski's Chat-Tyrant, Silvicultrix jelskii
- Golden-browed Chat-Tyrant, Silvicultrix pulchella
- Patagonian Tyrant, Colorhamphus parvirostris
- Tumbes Tyrant, Ochthoeca salvini
- Maroon-belted Chat-Tyrant, Ochthoeca thoracica
- Brown-backed Chat-Tyrant, Ochthoeca fumicolor
- Slaty-backed Chat-Tyrant, Ochthoeca cinnamomeiventris
- Blackish Chat-Tyrant, Ochthoeca nigrita
- Rufous-breasted Chat-Tyrant, Ochthoeca rufipectoralis
- Piura Chat-Tyrant, Ochthoeca piurae
- White-browed Chat-Tyrant, Ochthoeca leucophrys
- d'Orbigny's Chat-Tyrant, Ochthoeca oenanthoides
Fluvicolini: Fluvicoline Flycatchers Swainson, 1832-33
- White-bearded Flycatcher, Phelpsia inornata
- Chapada Flycatcher, Guyramemua affine
- Southern Scrub-Flycatcher, Sublegatus modestus
- Northern Scrub-Flycatcher, Sublegatus arenarum
- Amazonian Scrub-Flycatcher, Sublegatus obscurior
- Long-tailed Tyrant, Colonia colonus
- White-headed Marsh Tyrant, Arundinicola leucocephala
- Masked Water-Tyrant, Fluvicola nengeta
- Pied Water-Tyrant, Fluvicola pica
- Black-backed Water-Tyrant, Fluvicola albiventer
- Vermilion Flycatcher, Pyrocephalus obscurus
- Scarlet Flycatcher, Pyrocephalus rubinus
- Darwin's Flycatcher, Pyrocephalus nanus
- †San Cristobal Flycatcher, Pyrocephalus dubius
- Shear-tailed Gray Tyrant, Muscipipra vetula
- Streamer-tailed Tyrant, Gubernetes yetapa
- Black-and-white Monjita, Heteroxolmis dominicanus
- Cock-tailed Tyrant, Alectrurus tricolor
- Strange-tailed Tyrant, Alectrurus risora
Contopini: Pewees, Phoebes, Empids Fitzpatrick, 2004
- Drab Water Tyrant, Ochthornis littoralis
- Fuscous Flycatcher, Cnemotriccus fuscatus
- Tawny-chested Flycatcher, Aphanotriccus capitalis
- Black-billed Flycatcher, Aphanotriccus audax
- Euler's Flycatcher, Lathrotriccus euleri
- Gray-breasted Flycatcher, Lathrotriccus griseipectus
- Pileated Flycatcher, Xenotriccus mexicanus
- Belted Flycatcher, Xenotriccus callizonus
- Say's Phoebe, Sayornis saya
- Eastern Phoebe, Sayornis phoebe
- Black Phoebe, Sayornis nigricans
- Acadian Flycatcher, Empidonax virescens
- White-throated Flycatcher, Empidonax albigularis
- Alder Flycatcher, Empidonax alnorum
- Willow Flycatcher, Empidonax traillii
- Yellow-bellied Flycatcher, Empidonax flaviventris
- Yellowish Flycatcher, Empidonax flavescens
- Pacific-slope Flycatcher, Empidonax difficilis
- Cordilleran Flycatcher, Empidonax occidentalis
- Hammond's Flycatcher, Empidonax hammondii
- Dusky Flycatcher / American Dusky Flycatcher, Empidonax oberholseri
- Pine Flycatcher, Empidonax affinis
- Least Flycatcher, Empidonax minimus
- Gray Flycatcher / American Gray Flycatcher, Empidonax wrightii
- Buff-breasted Flycatcher, Empidonax fulvifrons
- Black-capped Flycatcher, Empidonax atriceps
- Olive Flycatcher / Olive Tufted-Flycatcher, Mitrephanes olivaceus
- Mexican Tufted-Flycatcher, Mitrephanes phaeocercus
- Costa Rican Tufted-Flycatcher, Mitrephanes aurantiiventris
- Choco Tufted-Flycatcher, Mitrephanes berlepschi
- Olive-sided Flycatcher, Contopus cooperi
- Ochraceous Pewee, Contopus ochraceus
- Greater Pewee, Contopus pertinax
- Dark Pewee, Contopus lugubris
- Smoke-colored Pewee, Contopus fumigatus
- Jamaican Pewee, Contopus pallidus
- Tumbes Pewee, Contopus punensis
- White-throated Pewee, Contopus albogularis
- Blackish Pewee, Contopus nigrescens
- Southern Tropical Pewee, Contopus cinereus
- Western Wood-Pewee, Contopus sordidulus
- Northern Tropical Pewee, Contopus bogotensis
- Eastern Wood-Pewee, Contopus virens
- Lesser Antillean Pewee, Contopus latirostris
- Cuban Pewee, Contopus caribaeus
- Hispaniolan Pewee, Contopus hispaniolensis
Taeniopterini: Monjitas, and Ground, Black, and Bush Tyrants Bonaparte, 1838
- Yellow-browed Tyrant, Satrapa icterophrys
- Little Ground-Tyrant, Syrtidicola fluviatilis
- Spot-billed Ground-Tyrant, Muscisaxicola maculirostris
- White-fronted Ground-Tyrant, Muscisaxicola albifrons
- Ochre-naped Ground-Tyrant, Muscisaxicola flavinucha
- Plain-capped Ground-Tyrant / Paramo Ground-Tyrant, Muscisaxicola alpinus
- Taczanowski's Ground-Tyrant, Muscisaxicola griseus
- Rufous-naped Ground-Tyrant, Muscisaxicola rufivertex
- Cinereous Ground-Tyrant, Muscisaxicola cinereus
- Dark-faced Ground-Tyrant, Muscisaxicola maclovianus
- White-browed Ground-Tyrant, Muscisaxicola albilora
- Cinnamon-bellied Ground-Tyrant, Muscisaxicola capistratus
- Puna Ground-Tyrant, Muscisaxicola juninensis
- Black-fronted Ground-Tyrant, Muscisaxicola frontalis
- Andean Negrito, Lessonia oreas
- Austral Negrito, Lessonia rufa
- Spectacled Tyrant, Hymenops perspicillatus
- Blue-billed Black-Tyrant, Knipolegus cyanirostris
- Plumbeous Tyrant, Knipolegus cabanisi
- Jelski's Black-Tyrant, Knipolegus signatus
- White-rumped Black-Tyrant, Knipolegus heterogyna
- Cinereous Tyrant, Knipolegus striaticeps
- White-winged Black-Tyrant, Knipolegus aterrimus
- Hudson's Black-Tyrant, Knipolegus hudsoni
- Rufous-tailed Tyrant, Knipolegus poecilurus
- Amazonian Black-Tyrant, Knipolegus poecilocercus
- Riverside Tyrant, Knipolegus orenocensis
- Crested Black-Tyrant, Knipolegus lophotes
- Caatinga Black-Tyrant / Sao Francisco Black-Tyrant, Knipolegus franciscanus
- Velvety Black-Tyrant, Knipolegus nigerrimus
- Red-rumped Bush-Tyrant, Cnemarchus erythropygius
- Rufous-webbed Bush-Tyrant, Polioxolmis rufipennis
- White-rumped Monjita, Xolmis velatus
- White Monjita, Xolmis irupero
- Fire-eyed Diucon, Pyrope pyrope
- Gray Monjita, Nengetus cinereus
- Black-crowned Monjita, Neoxolmis coronatus
- Chocolate-vented Tyrant, Neoxolmis rufiventris
- Salinas Monjita, Neoxolmis salinarum
- Rusty-backed Monjita, Neoxolmis rubetra
- Streak-throated Bush-Tyrant, Myiotheretes striaticollis
- Rufous-bellied Bush-Tyrant, Myiotheretes fuscorufus
- Santa Marta Bush-Tyrant, Myiotheretes pernix
- Smoky Bush-Tyrant, Myiotheretes fumigatus
- Black-billed Shrike-Tyrant, Agriornis montanus
- Lesser Shrike-Tyrant, Agriornis murinus
- White-tailed Shrike-Tyrant, Agriornis albicauda
- Gray-bellied Shrike-Tyrant, Agriornis micropterus
- Great Shrike-Tyrant, Agriornis lividus