Swallows
The swallows form an independent lineage in Sylvioidea tree. After the swallows, the remaining Sylvioidea species fall into two groups. The first include the Phylloscopidae leaf warblers, the other includes the bulbuls, babblers, and Sylvia warblers.
Hirundinidae: Martins & Swallows Rafinesque, 1815
21 genera, 92 species HBW-9
The organization of the swallows is now based on Schield et al. (2024), with some help from Brown (2019), de Silva et al. (2018), A.E. Johnson et al. (2016), and Sheldon et al. (2005).
The swallows split from the remaining Sylvioidea around 20 mya. Oliveros et al., (2019) put the split at 22 mya; Kuhl et al., (2021) have it at 22 mya; Stiller et al. (2024) say 17.3 mya. As there are nearly 100 species involved, I've inserted subfamiles and tribes to make the organization a bit clearer. The first split is the river martins (Pseudochelidoninae), followed by a split between the sawwings (Psalidoprocninae) and everything else (Hirundinae). Hirundinae divides into two groups, one that builds nests from mud (Hirundinini) and the other that nests in burrows and cavities (Prognini). The genus tree below shows this in more detail. You can also click for species tree.
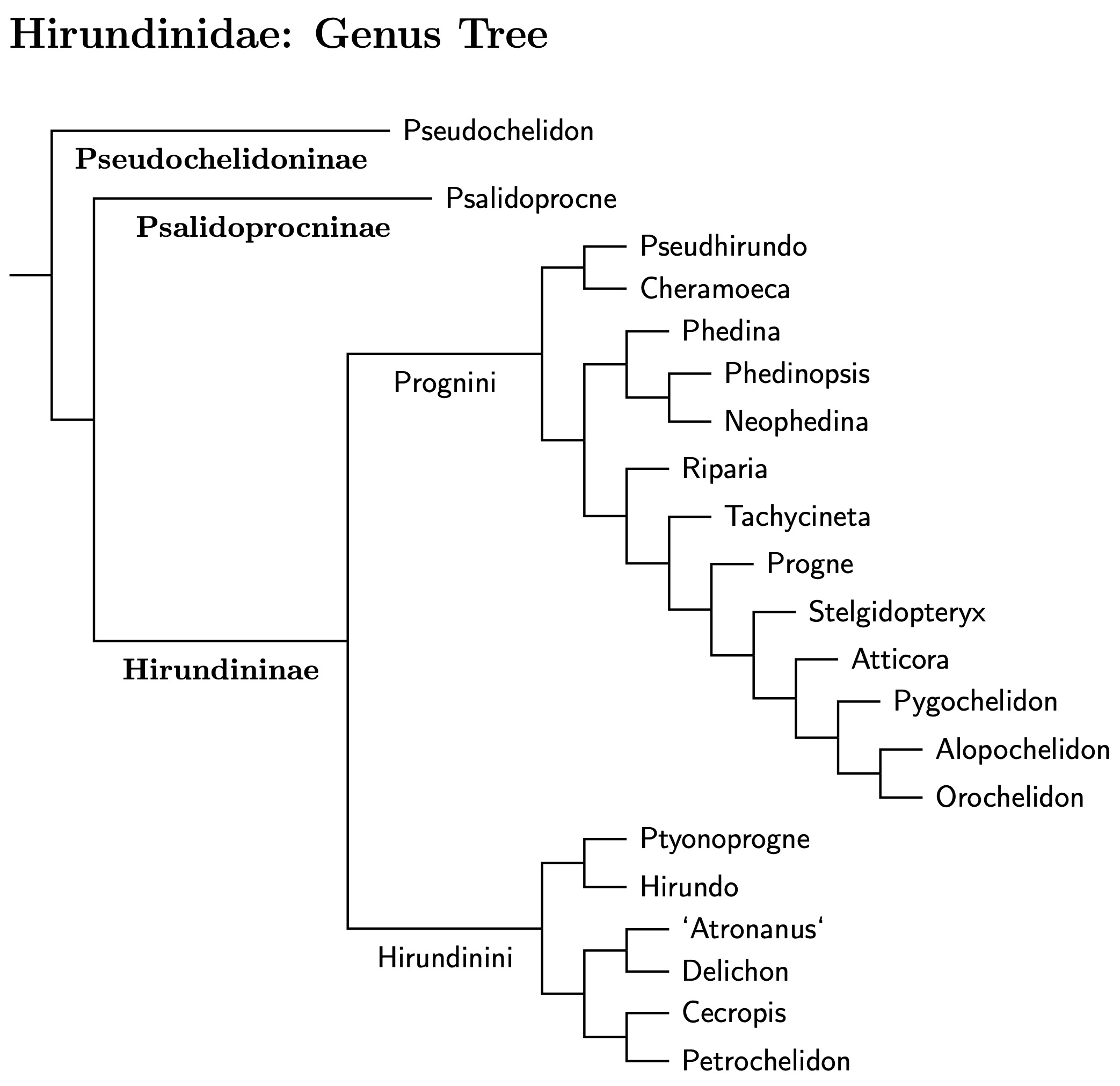 |
| Click diagram for Hirundinidae species tree |
|---|
Older Changes: The Rough-winged Swallows (genus Stelgidopteryx) are treated here as two species, but there is substantial genetic distance within both species (Babin, 2005), and they may be better treated as several species. Although "Ridgway's Swallow", Stelgidopteryx ridgwayi, is relatively distant from the Northern Rough-winged Swallow subspecies serripennis and fulvipennis, this distance is within the range of both the Southern Rough-winged Swallow and the remaining races of Northern Rough-winged Swallow. Further research, including more comprehensive genetic testing of the various races and at more locations, will be necessary to sort this out.
Common House-Martin Split: Common House-Martin, Delichon urbicum, is split into Western House-Martin, Delichon urbicum, and Siberian House-Martin, Delichon lagopodum. See Leader et al. (2021).
Cave Swallow: The Cave Swallow, Petrochelidon fulva, which includes two geographically-based distinct color morphs, is sometimes considered a candidate for a split. However, Kirchman et al. (2000) found substantial gene flow between the various populations. It was also striking how genetically distinct the Florida population had become in just a few years.
2025 Genus changes: De Silva et al. (2018) discovered that the Forest Swallow, formerly in genus Petrochelidon, was actually sister to the Delichon swallows. They attempted to establish the new genus ‘Atronanus’ for it, but didn't preregister it with ZooBank. As a result, I'm referring to the Forest Swallow as ‘Atronanus’ fuliginosus.
H&M-4 had already split Phedina into 3 monotypic genera. This is consistent with Schield et al. (2024). As a result, the Banded Martin, Phedina cincta, becomes Neophedina cincta (Roberts 1922) and Brazza's Martin, Phedina brazzae, becomes Phedinopsis brazzae (Wolters 1971).
2025 Species changes: The Rock Martin, Ptyonoprogne fuligula has been split into Large Rock Martin, Ptyonoprogne fuligula, and Red-throated Rock Martin, Ptyonoprogne rufigula. The Large Rock Martin consists of the more southern subspecies anderssoni, fuligula, and pretoriae. The Red-throated Rock Martin has the northern subspecies pusilla, bansoensis and rufigula. See Brown (2019).
The Pacific Swallow, Hirundo tahitica, has been split into Tahiti Swallow, Hirundo tahitica and Pacific Swallow, Hirundo javanica based on morphological differences (HBW/BirdLife).
There are a complex set of changes to the Red-rumped Swallows (Cecropis).
- The monotypic European Red-rumped Swallow, Cecropis rufula, is split from Red-rumped Swallow, Cecropis daurica.
- Three subspecies of Red-rumped Swallow, melanocrissus, kumboensis, and emini are merged with the West African Swallow Cecropis domicella. Both the commmon and scientific names change, giving us African Red-rumped Swallow, Cecropis melanocrissus.
- The remaining subspecies of Red-rumped Swallow, Cecropis daurica: daurica, japonica, nipalensis, and erythropygia join with all four subspecies of Striated Swallow, Cecropis striolata, to create the Eastern Red-rumped Swallow, Cecropis daurica. The name daurica has priority over striolata.
Finally, the Madagascan Martin, Riparia cowani, has been split from the Brown-throated Martin Riparia paludicola. Shield et al. (2024) found they split a healthy 3.3 million years ago. BoW and HBW/BirdLife support the split based on morphology and vocalizations.
There has also been some rearrangement of the linear order. Here's the result.
Pseudochelidoninae: River Martins Shelly, 1896
- African River-Martin, Pseudochelidon eurystomina
- White-eyed River-Martin, Pseudochelidon sirintarae
Psalidoprocninae: Sawwings Sharpe, 1870
- Square-tailed Sawwing, Psalidoprocne nitens
- Fanti Sawwing, Psalidoprocne obscura
- Mountain Sawwing, Psalidoprocne fuliginosa
- Black Sawwing, Psalidoprocne pristoptera
- White-headed Sawwing, Psalidoprocne albiceps
Hirundininae Rafinesque, 1815
Prognini: Martins & Allies Cassin, 1853
- Gray-rumped Swallow, Pseudhirundo griseopyga
- White-backed Swallow, Cheramoeca leucosterna
- Mascarene Martin, Phedina borbonica
- Banded Martin, Phedina cincta
- Brazza's Martin, Phedina brazzae
- Pale Martin, Riparia diluta
- Bank Swallow / Sand Martin, Riparia riparia
- Congo Martin, Riparia congica
- Gray-throated Martin, Riparia chinensis
- Brown-throated Martin, Riparia paludicola
- Madagascan Martin, Riparia cowani
- Tree Swallow, Tachycineta bicolor
- Golden Swallow, Tachycineta euchrysea
- Violet-green Swallow, Tachycineta thalassina
- Bahama Swallow, Tachycineta cyaneoviridis
- Tumbes Swallow, Tachycineta stolzmanni
- Mangrove Swallow, Tachycineta albilinea
- White-winged Swallow, Tachycineta albiventer
- White-rumped Swallow, Tachycineta leucorrhoa
- Chilean Swallow, Tachycineta leucopyga
- Brown-chested Martin, Progne tapera
- Peruvian Martin, Progne murphyi
- Purple Martin, Progne subis
- Southern Martin, Progne elegans
- Gray-breasted Martin, Progne chalybea
- Galapagos Martin, Progne modesta
- Sinaloa Martin, Progne sinaloae
- Cuban Martin, Progne cryptoleuca
- Caribbean Martin, Progne dominicensis
- Northern Rough-winged Swallow, Stelgidopteryx serripennis
- Southern Rough-winged Swallow, Stelgidopteryx ruficollis
- White-banded Swallow, Atticora fasciata
- Black-capped Swallow, Atticora pileata
- White-thighed Swallow, Atticora tibialis
- Blue-and-white Swallow, Pygochelidon cyanoleuca
- Black-collared Swallow, Pygochelidon melanoleuca
- Tawny-headed Swallow, Alopochelidon fucata
- Pale-footed Swallow, Orochelidon flavipes
- Brown-bellied Swallow, Orochelidon murina
- Andean Swallow, Orochelidon andecola
Hirundinini: Mud-nesting Swallows Rafinesque, 1815
- Large Rock Martin, Ptyonoprogne fuligula
- Dusky Crag-Martin, Ptyonoprogne concolor
- Eurasian Crag-Martin, Ptyonoprogne rupestris
- Pale Crag-Martin, Ptyonoprogne obsoleta
- Red-throated Rock Martin, Ptyonoprogne rufigula
- Black-and-rufous Swallow, Hirundo nigrorufa
- Blue Swallow, Hirundo atrocaerulea
- Pied-winged Swallow, Hirundo leucosoma
- White-tailed Swallow, Hirundo megaensis
- Pearl-breasted Swallow, Hirundo dimidiata
- Hill Swallow, Hirundo domicola
- Pacific Swallow, Hirundo javanica
- Welcome Swallow, Hirundo neoxena
- Tahiti Swallow, Hirundo tahitica
- Wire-tailed Swallow, Hirundo smithii
- White-throated Swallow, Hirundo albigularis
- White-bibbed Swallow, Hirundo nigrita
- Barn Swallow, Hirundo rustica
- Ethiopian Swallow, Hirundo aethiopica
- Red-chested Swallow, Hirundo lucida
- Angolan Swallow, Hirundo angolensis
- Forest Swallow, ‘Atronanus’ fuliginosa
- Western House-Martin, Delichon urbicum
- Siberian House Martin, Delichon lagopodum
- Asian House-Martin, Delichon dasypus
- Nepal House-Martin, Delichon nipalense
- Lesser Striped-Swallow, Cecropis abyssinica
- Red-breasted Swallow, Cecropis semirufa
- Greater Striped-Swallow, Cecropis cucullata
- Mosque Swallow, Cecropis senegalensis
- Rufous-bellied Swallow, Cecropis badia
- African Red-rumped Swallow, Cecropis melanocrissus
- European Red-rumped Swallow, Cecropis rufula
- Eastern Red-rumped Swallow, Cecropis daurica
- Sri Lanka Swallow, Cecropis hyperythra
- South African Swallow / South African Cliff-Swallow, Petrochelidon spilodera
- Preuss's Swallow / Preuss's Cliff-Swallow, Petrochelidon preussi
- Red-throated Swallow / Red-throated Cliff-Swallow, Petrochelidon rufigula
- Red Sea Swallow / Red Sea Cliff-Swallow, Petrochelidon perdita
- Tree Martin, Petrochelidon nigricans
- Streak-throated Swallow, Petrochelidon fluvicola
- Fairy Martin, Petrochelidon ariel
- Cliff Swallow / American Cliff-Swallow, Petrochelidon pyrrhonota
- Cave Swallow, Petrochelidon fulva
- Chestnut-collared Swallow, Petrochelidon rufocollaris
Hyliidae, Aegithalidae, Cettiidae, and Phylloscopidae
These four families are relatively closely related.
Hyliidae: Hylias Bannerman, 1923
2 genera, 2 species Not HBW Family
- Green Hylia, Hylia prasina
- Tit Hylia, Pholidornis rushiae
I orginally put the Hylias in a separate family based on Johanssen et al. (2008). Fregin et al. (2012) place them sister to the long-tailed tits, albeit with less support than one might like. That seems to be not quite correct as Oliveros et al. (2019), Kuhl et al. (2021), and Stiller et al. (2024) place them sister to the combined long-tailed tits and Cettiidae. Estimates of the most recent common ancestor of the tree families include 20 mya (Oliveros et al.), 17.8 mya (Kuhl et al.), and 17.5 mya (Stiller et al.). Note that all three call Cettiidae “Scotocercidae”, which is incorrect. See below in the Cettiidae section.
Classification of the Tit-Hylia (Pholidornis rushiae) has long been controversial. It has variously been placed in at least 7 other families: Sylviidae, Estrildidae, Dicaeidae, Nectariniidae, Remizidae, Hyliidae and Meliphagidae. Sefc et al. (2003) found Hylia and Pholidornis more closely related than either is to Aegithalos or Phylloscopus. More recently, Oliveros et al. (2019) considered them sister taxa. with their most recent common ancestor living about 15 mya.
Aegithalidae: Long-tailed Tits Reichenbach, 1849-50
3 genera, 13 species HBW-13
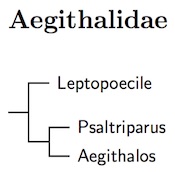 |
| Click for Aegithalidae tree |
|---|
Leptopoecile has been included in Aegithalidae rather than Sylviidae based on Sturmbauer et al. (1998), Alström et al. (2006), Johansson et al. (2008b), and Päckert et al. (2010).
The current arrangement of Aegithalos is based on the results of Päckert et al. (2010). However, they found that the last four species A. bonvaloti through A. sharpei are extremely close genetically, close enough to call into serious question whether they are separate species. Nonetheless, they appear distinct and, according to HBW-13, there is no evidence of interbreeding in areas of overlap. Päckert et al. also found quite large genetic differences between the various races of A. concinnus, suggesting several species are involved. This had previously been suspected based on plumage, but the presence of zones of intergradation has kept them lumped together. Unfortunately, Päckert et al. did not examine the entire concinnus complex, so the situation is not entirely clear, even without the complication of intergrades. Until more information is available, I leave the species limits unchanged.
Johansson et al. (2016) found that Psaltria is embedded in the concinnus complex (which still needs to be sorted out). I have merged Psaltria into Aegithalos.
- White-browed Tit-warbler, Leptopoecile sophiae
- Crested Tit-warbler, Leptopoecile elegans
- Bushtit / American Bushtit, Psaltriparus minimus
- White-cheeked Tit / White-cheeked Bushtit, Aegithalos leucogenys
- Black-throated Tit / Black-throated Bushtit, Aegithalos concinnus
- Pygmy Tit / Pygmy Bushtit, Aegithalos exilis
- Long-tailed Tit, Aegithalos caudatus
- Silver-throated Tit / Silver-throated Bushtit, Aegithalos glaucogularis
- White-throated Tit / White-throated Bushtit, Aegithalos niveogularis
- Black-browed Tit / Black-browed Bushtit, Aegithalos bonvaloti
- Sooty Tit / Sooty Bushtit, Aegithalos fuliginosus
- Rufous-fronted Tit / Rufous-fronted Bushtit, Aegithalos iouschistos
- Burmese Tit / Burmese Bushtit, Aegithalos sharpei
Cettiidae: Cettiid Warblers Coues, 1903
12 genera, 36 species Not HBW Family
The name Cettiidae: Note that Cettiidae is incorrectly called Scotocercidae by all in the big three papers. They were apparently unaware that Cettiinae was used as a family-group name by Coues, 1903, making it available for use at family level. It's first use at the family level (Cettiidae) may have been by Chigi in 1912. It was in recent use until after Scotocercidae was invented by Fregin et al. (2012). Since Cettiidae has more than a century of priority over Scotocercidae, it is the correct name for this famiy. I thank Laurent Raty for pointing this out on BIrdForum.
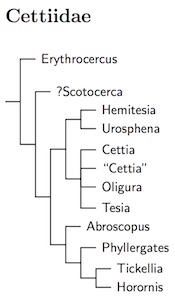 |
| Click for Cettiidae tree |
|---|
We briefly moved back to familiar territory family-wise, with the swallows and then the long-tailed tits. The long-tailed tits seem to be the sister group of another new family, the cettiid bush-warblers (Cettiidae). This group has an primarily south and east Asian distribution ranging into Oceania, the Western Palearctic, and tropical Africa. Although IOC and Clements use the term “Bush-Warblers and allies” for the family, I think it's a problem because there are as many “Bush-Warblers” in Locustellidae as in Cettiidae.
Fregin et al. (2012) proposed treating the genus Erythrocercus as a separate family, Erythrocercidae. One reason for separating them is that they have 12 rectrices rather than the 10 which the other Cettiidae have. Further, they are African rather than Asian (only one purely African species would be left in Cettiidae), and they are a fairly deep branch in the Sylvioidea tree. However, I am not yet persuaded we need another tiny family here. Since I wrote that, Oliveros et al. (2019) estimated the split between Erythrocercus and the rest of Cettiidae occurred about 17.8 mya. More recently, Kuhl et al. (2021) estimated the split at 14.4 mya and Stiller et al. (2024) put it a little earlier, at 13.4 mya. That's rather young for a family in this part of the avian tree, even if they do have 2 extra tail feathers. It is perfect for ranking them as a subfamily, Erythrocercinae.
That automatically means that Scotocerca should not be raised to family status. Both are treated as subfamilies here. For the record, Oliveros et al. put the split of Scotocerca at just over 15 mya.
There had once been questions about which family Scotocerca inquieta belongs to. It was often been placed with the cisticolas, but Barhoum and Burns (2002) presented evidence suggesting that Scotocerca is part of Acrocephalidae, not Cisticolidae. However, there were issues about the sample used, and the result did not appear robust. In truth, it was also wrong. Alström et al. (2011a) have carried out a more comprehensive analysis and found that Scotocerca is actually a cettiid warbler. They found it sister to a clade containing Cettia and Abroscopus. More recently Oliveros et al. (2019) has backed this up.
Irestedt et al. (2011) found that Neumann's Warbler, Hemitesia neumanni, is a member of the Cettiidae. It is most likely closest to the Urosphena stubtails, although the relationship is somewhat distant. Given that most of the Cettiidae are oriental, it's interesting that the African genus ended up among them. Irestedt et al. (2011) also provided stronger support for the basal position of Erythrocercus previously noted by Johansson et al. (2008b). Since Erythrocercus is also African, this opens the possibility that the Cettiidae may have originated in Africa.
The Phyllergates tailorbirds were formerly placed among the cisticolas in the genus Orthotomus (see Alström et al., 2006, 2011d; Fuchs et al., 2006a). The Odedi Bush-Warbler is a recent discovery (LeCroy and Barker, 2006). Its song had been heard by Jared Diamond in 1972, but the bird was not tracked down until recently. Hadden was able to mist-net the bird in 2000.
The arrangement of genera is now based on Alström et al. (2011d). This is generally consistent with my previous treatment based on Alström et al. (2006), Fuchs et al. (2006a), Johansson et al. (2008b), and Irestedt et al. (2011).
The genus Cettia as usually constituted is polyphyletic. Although my original guess was that it involved two different groups, Alström et al. (2011d) found that the situation was more complex. Accordingly, I've transferred the Pale-footed Bush-Warbler to Hemitesia, Gray-sided Bush-Warbler to Oligura, and temporarily designated Chestnut-crowned Bush-Warbler as “Cettia”. Only Cetti's Warbler is left in Cettia (as in the previous TiF version). The other former Cettia warblers are placed in Horornis (Hodgson 1845, type fortipes). I had previously put them in Horeites in spite of some uncertainty about whether the type species, brunnifrons, actually belonged with them. Well, it doesn't.
Alström et al. (2011d) suggest a somewhat different set of generic limits, with Hemitesia merged into Urosphena, and with Cettia including “Cettia” and Oligura. I decided to not follow that because (1) the genera I use seem to mark relatively deep divisions, and (2) there is some uncertainty about whether such a broad Cettia would also have to include Tesia (there's a reason I have a 4-fold polytomy there; see Fig. 3 and the supplementary material in Alström et al., 2011d).
Finally, the Manchurian Bush-Warbler is restricted to the race borealis, with canturians being demoted to a subspecies of Japanese Bush-Warbler, Horornis diphone. Genetically, borealis is closer to the Philippine Bush-Warbler than to the Japanese Bush-Warbler races cantans canturians, justifying treatment as a separate species. As borealis breeds in Manchuria, while canturians does not, I've retained the name Manchurian Bush-Warbler. Since canturians is quite close to cantans, and presumably to diphone, it is treated as a subspecies of the Japanese Bush-Warbler. That said, diphone itself has not been studied, and there are differences (e.g., song) that suggest canturians may be a separate biological species from cantans and diphone. A more comprehensive look at this complex would be helpful (including restricta).
Erythrocercinae: Bristle-flycatchers Fregin et al., 2012
- Little Yellow Flycatcher, Erythrocercus holochlorus
- Chestnut-capped Flycatcher, Erythrocercus mccallii
- Livingstone's Flycatcher, Erythrocercus livingstonei
Scotocercinae: Streaked Scrub Warbler Fregin et al., 2012
- Streaked Scrub Warbler, Scotocerca inquieta
Cettiinae: Cettiid Warblers Alström, Olsson, & Ericson, 2014
- Neumann's Warbler, Hemitesia neumanni
- Pale-footed Bush-Warbler, Hemitesia pallidipes
- Asian Stubtail, Urosphena squameiceps
- Bornean Stubtail, Urosphena whiteheadi
- Timor Stubtail, Urosphena subulata
- Cetti's Warbler, Cettia cetti
- Chestnut-crowned Bush-Warbler, "Cettia" major
- Gray-sided Bush-Warbler, Oligura brunnifrons
- Chestnut-headed Tesia, Oligura castaneocoronata
- Gray-bellied Tesia, Tesia cyaniventer
- Slaty-bellied Tesia, Tesia olivea
- Javan Tesia, Tesia superciliaris
- Russet-capped Tesia, Tesia everetti
- Yellow-bellied Warbler, Abroscopus superciliaris
- Black-faced Warbler, Abroscopus schisticeps
- Rufous-faced Warbler, Abroscopus albogularis
- Mountain Tailorbird, Phyllergates cuculatus
- Rufous-headed Tailorbird, Phyllergates heterolaemus
- Broad-billed Warbler, Tickellia hodgsoni
- Brown-flanked Bush-Warbler, Horornis fortipes
- Aberrant Bush-Warbler, Horornis flavolivaceus
- Sunda Bush-Warbler, Horornis vulcanius
- Hume's Bush-Warbler, Horornis brunnescens
- Yellow-bellied Bush-Warbler, Horornis acanthizoides
- Manchurian Bush-Warbler, Horornis borealis
- Philippine Bush-Warbler, Horornis seebohmi
- Japanese Bush-Warbler, Horornis diphone
- Palau Bush-Warbler, Horornis annae
- Tanimbar Bush-Warbler, Horornis carolinae
- Shade Bush-Warbler, Horornis parens
- Odedi / Bougainville Bush-Warbler, Horornis haddeni
- Fiji Bush-Warbler, Horornis ruficapilla
Phylloscopidae: Leaf-Warblers Jerdon, 1863 (1854)
9 genera, 79 species Not HBW Family
These are followed by the leaf-warblers (Phylloscopidae), which is a new family carved out of Sylviidae. It includes about 80 species, now generally classified in a single genus Phylloscopus. The genus Seicercus was previously recognized, but DNA made the two genus treatment untenable. There needed to be more as in TiF, or just one.
The taxonomy here is primarily based on Johansson et al. (2007a) and Olsson et al. (2005), with Irwin et al. (2005) and Martens et al. (2008) filling in some details. One way to handle the resulting changes is to move many species from Phylloscopus to Seicercus. This approach was previously taken in the TiF checklist. However, I think it is better to recognize the major clades as genera. Every genus recognized differs in DNA by at least 12%, and is visually distinct from its nearest relative. Some subclades that also differ by 12% or more, but are not visually distinct, are not recognized as separate genera.
Both Johansson et al. (2007a) and Olsson et al. (2005) are consistent with a division into nine clades, each of which I recognize as a separate genus. A brief description of the typical member of each genus is given below. Some members of the various genera may differ from the description. As is commonly done, I use the term “phylloscops” to describe any of the eye-lined warblers typically placed in the genus Phylloscopus.
- Abrornis. Kinglet-like phylloscops: maculipennis through proregulus;
- Rhadina. Plain phylloscops with pale-edged tertials: orientalis, bonelli, and sibilatrix;
- Phylloscopus. Plain phylloscops without wingbars: tytleri through canariensis;
- Pindalus. Reddish phylloscops: umbrovirens through laetus;
- Seicercus. Spectacled warblers: poliogenys through soror;
- Pycnosphrys: Reddish spectacled warblers: castaniceps, montis, and grammiceps;
- "Pycnosphrys": Fairly plain phylloscops, possibly with wingbars: cebuensis to ijimae;
- Acanthopneuste: Phylloscops with wingbars: xanthodryas through plumbeitarsus;
- Cryptigata: Capped phylloscops: ricketti to the end.
They also agree that clades 8 and 9 are sisters, as are (1)-(3) and (4)-(9). Olsson et al. (2004) has yet another topology, but the same clades appear. Previously, clades (1)-(4), and (7)-(9) were considered Phylloscopus while (5) and (6) were considered Seicercus.
I've split Japanese Leaf-Warbler, Acanthopneuste xanthodryas, and Kamchatka Leaf-Warbler, Acanthopneuste examinandus, from Arctic Warbler, Acanthopneuste borealis. Reeves et al. (2008) found substantial genetic differences between the Kamchatka/Sakhalin populations and borealis/kennicotti. They suggested separating them as “Pacific Warbler”. However, Saitoh et al. (2010) also included Japanese populations of in their analysis. They found that the Hokkaido birds grouped with those on Sakhalin Island and in Kamchatcka, but that the other Japanese populations were well-separated from the rest. Additional races are sometimes recognized, but they appear to belong to the borealis/kennicotti clade (Alström et al., 2011c). The English names are those of Alström et al. (2011c), which contains further information on the split. Note that there is a record of Kamchatka Leaf-Warbler from Amchitka Island, Alaska (Kenyon, 1961).
I've handled the Acanthopneuste trochiloides complex in an unusual way. This includes nitidus, viridanus, trochiloides, obscuratus, plumbeitarsus and S. v. ludlowli (or at least part of it). Irwin et al. (2001) found two groups. The first comprised of nitidus and viridanus, the second including trochiloides, obscuratus, and plumbeitarsus in a trichotomy. Although ludlowli had its own clade, it was too closely related to viridis to separate. This gives us several choices: lump them all, split the first group (which are more distantly related) and lump the second, or split them all. I take the last option here.
In contrast, BLI and Clements lump them all, which is defensible. Sibley and Monroe, Dickinson, IOC, and HBW take a different route. They split trochiloides and plumbeitarsus. This is not consistent with Irwin et al.'s results. It conflicts with the status of obscuratus as an equal member of the trichotomy. It also includes taxa from both groups in trochiloides.
For information concerning the recently recognized Limestone Leaf-Warbler, Cryptigata calciatilis, see Alström et al. (2010).
- Buff-barred Warbler, Abrornis pulcher
Click for Phylloscopidae tree - Ashy-throated Warbler, Abrornis maculipennis
- Yellow-browed Warbler, Abrornis inornatus
- Hume's Leaf-Warbler, Abrornis humei
- Chinese Leaf-Warbler, Abrornis yunnanensis
- Brooks's Leaf-Warbler, Abrornis subviridis
- Lemon-rumped Warbler, Abrornis chloronotus
- Sichuan Leaf-Warbler, Abrornis forresti
- Gansu Leaf-Warbler, Abrornis kansuensis
- Pallas's Leaf-Warbler, Abrornis proregulus
- Eastern Bonelli's Warbler, Rhadina orientalis
- Western Bonelli's Warbler, Rhadina bonelli
- Wood Warbler, Rhadina sibilatrix
- Tytler's Leaf-Warbler, Phylloscopus tytleri
- Yellow-streaked Warbler, Phylloscopus armandii
- Radde's Warbler, Phylloscopus schwarzi
- Alpine Leaf-Warbler, Phylloscopus occisinensis
- Tickell's Leaf-Warbler, Phylloscopus affinis
- Sulphur-bellied Warbler, Phylloscopus griseolus
- Dusky Warbler, Phylloscopus fuscatus
- Smoky Warbler, Phylloscopus fuligiventer
- Buff-throated Warbler, Phylloscopus subaffinis
- Willow Warbler, Phylloscopus trochilus
- Mountain Chiffchaff, Phylloscopus sindianus
- Plain Leaf-Warbler, Phylloscopus neglectus
- Common Chiffchaff, Phylloscopus collybita
- Iberian Chiffchaff, Phylloscopus ibericus
- Canary Islands Chiffchaff, Phylloscopus canariensis
- Brown Woodland-Warbler, Pindalus umbrovirens
- Black-capped Woodland-Warbler, Pindalus herberti
- Uganda Woodland-Warbler, Pindalus budongoensis
- Yellow-throated Woodland-Warbler, Pindalus ruficapilla
- Laura's Woodland-Warbler, Pindalus laurae
- Red-faced Woodland-Warbler, Pindalus laetus
- Gray-cheeked Warbler, Seicercus poliogenys
- White-spectacled Warbler, Seicercus affinis
- Green-crowned Warbler, Seicercus burkii
- Gray-crowned Warbler, Seicercus tephrocephalus
- Whistler's Warbler, Seicercus whistleri
- Bianchi's Warbler, Seicercus valentini
- Martens's Warbler, Seicercus omeiensis
- Alstrom's Warbler, Seicercus soror
- Yellow-breasted Warbler, Pycnosphrys montis
- Chestnut-crowned Warbler, Pycnosphrys castaniceps
- Sunda Warbler, Pycnosphrys grammiceps
- Lemon-throated Leaf-Warbler, "Pycnosphrys" cebuensis
- Philippine Leaf-Warbler, "Pycnosphrys" olivaceus
- Eastern Crowned-Warbler, "Pycnosphrys" coronatus
- Ijima's Leaf-Warbler, "Pycnosphrys" ijimae
- Japanese Leaf-Warbler, Acanthopneuste xanthodryas
- Arctic Warbler, Acanthopneuste borealis
- Kamchatka Leaf-Warbler, Acanthopneuste examinandus
- Large-billed Leaf-Warbler, Acanthopneuste magnirostris
- Pale-legged Leaf-Warbler, Acanthopneuste tenellipes
- Sakhalin Leaf-Warbler, Acanthopneuste borealoides
- Emei Leaf-Warbler, Acanthopneuste emeiensis
- Green Warbler, Acanthopneuste nitidus
- Western Greenish Warbler, Acanthopneuste viridanus
- Greenish Warbler, Acanthopneuste trochiloides
- Dull-green Warbler, Acanthopneuste obscuratus
- Two-barred Warbler, Acanthopneuste plumbeitarsus
- Sulphur-breasted Leaf-Warbler, Cryptigata ricketti
- Limestone Leaf-Warbler, Cryptigata calciatilis
- Yellow-vented Warbler, Cryptigata cantator
- Western Crowned-Warbler, Cryptigata occipitalis
- Hartert's Leaf-Warbler, Cryptigata goodsoni
- Blyth's Leaf-Warbler, Cryptigata reguloides
- Claudia's Leaf-Warbler, Cryptigata claudiae
- Hainan Leaf-Warbler, Cryptigata hainanus
- Kloss's Leaf-Warbler, Cryptigata ogilviegranti
- Davison's Leaf-Warbler, Cryptigata davisoni
- Gray-hooded Warbler, Cryptigata xanthoschistos
- Mountain Leaf-Warbler, Cryptigata trivirgata
- Timor Leaf-Warbler, Cryptigata presbytes
- Negros Leaf-Warbler, Cryptigata nigrorum
- Sulawesi Leaf-Warbler, Cryptigata sarasinorum
- Island Leaf-Warbler, Cryptigata maforensis
- Makira Leaf-Warbler, Cryptigata makirensis
- Kolombangara Leaf-Warbler, Cryptigata amoena
Bulbuls
The bulbuls stand between the leaf warbler group and the warbler-babbler clade that ends the Sylvioidea. Stiller et al. (2024) estimate they split from the other Sylvioidea about 16.25 mya. Kuhl et al. (2021) put it at 20.0 mya, and Oliveros et al. at 21.2 mya.
Pycnonotidae: Bulbuls G.R. Gray, 1840
28 genera, 153 species HBW-10
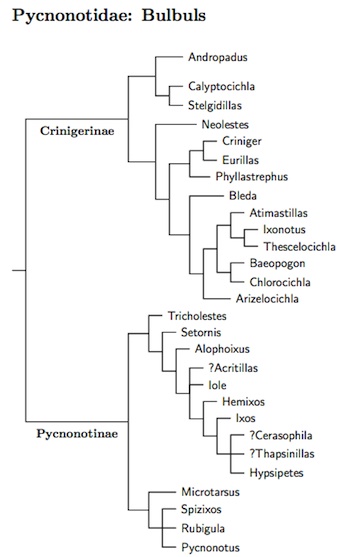 The bulbul family Pycnonotidae has previously seen some change in
composition and a reorganization.
The bulbul family Pycnonotidae has previously seen some change in
composition and a reorganization.
Genera formerly considered bulbuls include Nicator, now in its own family, Nicatoridae; Bernieria and Xanthomixis, now in Bernieridae; the vanga Tylas; and the dapplethroat Arcanator. Finally, Lioptilus belongs in Sylviidae. There had been some question whether Neolestes was a bulbul or not. Zuccon and Ericson (2010b) showed that it is a bulbul.
Pasquet et al. (2001) found two clades, the African greenbuls and the mostly Asian bulbuls. They also argued in favor of restructing some genera. The more recent papers by Moyle and Marks (2006) and Johansson et al. (2007b) sampled a large fraction of the species and permitted a comprehensive phyogenetic reorganization of the family. That said, the coverage of Pycnonotinae was less satisfactory. This has been partially remedied by Oliveros (2009) and Oliveros and Moyle (2010), which add more data on Philippine bulbuls to the Moyle and Marks (2006) data set. Zuccon and Ericson (2010b) included additional genetic data in their analysis.
Several genera have been reorganized. The formerly large genus Andropadus has been split into several 4 genera, including Stelgidillas, Eurillas, and Arizelocichla. Atimastillas is split from Chlorocichla. The Red-tailed Leaf-love (formerly Pyrrhurus scandens) is included in Phyllastrephus. Criniger has been split into the African Criniger and Asian Alophoixus.
The green-headed Montane Tiny-Greenbul, Phyllastrephus albigula, has been split from the gray-headed Lowland-Tiny Greenbul, Phyllastrephus debilis. See Fuchs et al. (2011a).
Based on Manawatthana et al. (2017), Charlotte's Bulbul, Iole charlottae (inc. perplexa) has been split from Buff-vented Bulbul, Iole olivacea, and Cachar Bulbul, Iole cacharensis, has been split from Olive Bulbul, Iole viridescens. Moreover, the subspecies lekhakuni and cinnamonomeoventris have been transferred from Gray-eyed Bulbul, Iole propinqua to Olive Bulbul, Iole viridescens while myitkyinensis has moved the other way, from Olive Bulbul to Gray-eyed Bulbul.
It's still not clear whether Pycnonotus is monophyletic (see HBW-10). However, it's getting closer. For some time, Pycnonotus has been overlumped. I've placed several former Pycnonotus species in Microtarsus, which appears as a basal group in Pycnonotinae (Moyle and Marks, 2006; Oliveros, 2009; Oliveros and Moyle, 2010). I had previously split part of Pycnonotus into Euptilosus, and Brachypodius, but it now seems clear they are better merged into Microtarsus. Finally, several species often placed in Pycnonotus are separated in the restored genus Rubigula.
I've also moved the Philippine members of Ixos into Hypsipetes, which has also absorbed Microscelis. The results of Oliveros (2009) and Oliveros and Moyle (2010) also allow an alternative treatment, placing the former Ixos in a separate genus. The Indian Ocean/Malagasy Hypsipetes have been rearranged in view of Warren et al. (2005). Further, the Moheli Bulbul, Hypsipetes moheliensis, has been split from the Comoros Bulbul (now Grand Comoro Bulbul), Hypsipetes parvirostris.
I have included the Bare-faced Bulbul, Pycnonotus hualon, described by Woxvold et al. (2009). Where exactly this goes in Pycnonotus is unclear, but after reading Woxvold et al. carefully, this is my best guess. As the authors note, Pycnonotus itself needs revision.
Oliveros (2009) and Oliveros and Moyle (2010) note several subspecies that appear to deserve species rank. Two species contain two or more separate clades. The IOC has provided English names for Ashy-fronted Bulbul, Pycnonotus cinereifrons (split from Olive-winged Bulbul, Pycnonotus plumosus), Palawan Bulbul, Alophoixus frater (split from Gray-cheeked Bulbul, Alophoixus bres), Visayan Bulbul, Hypsipetes guimarasensis, and Mindoro Bulbul, Hypsipetes mindorensis (both split from Philippine Bulbul, Hypsipetes philippinus). In all 4 cases, I'm presuming that each of the new species are monotypic. Further, there are indications that Pycnonotus urostictus atricaudatus, and one or more of the subspecies of Pycnonotus goiavier and Hypsipetes amaurotis should be considered distinct species, but these cases involve additional complexities.
Aceh Bulbul, Pycnonotus snouckaerti, has been split from Orange-spotted Bulbul, Pycnonotus bimaculatus, based on Eaton & Collar (2015).
Crinigerinae: Greenbuls Bonaparte, 1854 (1831)
- Sombre Greenbul, Andropadus importunus
- Golden Greenbul, Calyptocichla serinus
- Slender-billed Greenbul, Stelgidillas gracilirostris
- Black-collared Bulbul, Neolestes torquatus
- Western Bearded-Greenbul, Criniger barbatus
- Eastern Bearded-Greenbul, Criniger chloronotus
- Red-tailed Greenbul, Criniger calurus
- White-bearded Greenbul, Criniger ndussumensis
- Yellow-bearded Greenbul, Criniger olivaceus
- Yellow-whiskered Greenbul, Eurillas latirostris
- Little Greenbul, Eurillas virens
- Plain Greenbul, Eurillas curvirostris
- Little Gray Greenbul, Eurillas gracilis
- Ansorge's Greenbul, Eurillas ansorgei
- Red-tailed Leaf-love, Phyllastrephus scandens
- Terrestrial Brownbul, Phyllastrephus terrestris
- Northern Brownbul, Phyllastrephus strepitans
- Gray-olive Greenbul, Phyllastrephus cerviniventris
- Pale-olive Greenbul, Phyllastrephus fulviventris
- Baumann's Olive-Greenbul, Phyllastrephus baumanni
- Toro Olive-Greenbul, Phyllastrephus hypochloris
- Fischer's Greenbul, Phyllastrephus fischeri
- Cabanis's Greenbul, Phyllastrephus cabanisi
- Placid Greenbul, Phyllastrephus placidus
- Cameroon Olive-Greenbul, Phyllastrephus poensis
- Icterine Greenbul, Phyllastrephus icterinus
- Xavier's Greenbul, Phyllastrephus xavieri
- Liberian Greenbul, Phyllastrephus leucolepis
- Sassi's Olive-Greenbul, Phyllastrephus lorenzi
- White-throated Greenbul, Phyllastrephus albigularis
- Yellow-streaked Greenbul, Phyllastrephus flavostriatus
- Sharpe's Greenbul, Phyllastrephus alfredi
- Gray-headed Greenbul, Phyllastrephus poliocephalus
- Montane Tiny-Greenbul, Phyllastrephus albigula
- Lowland Tiny-Greenbul, Phyllastrephus debilis
- Red-tailed Bristlebill, Bleda syndactylus
- Green-tailed Bristlebill, Bleda eximius
- Yellow-lored Bristlebill, Bleda notatus
- Gray-headed Bristlebill, Bleda canicapillus
- Yellow-throated Leaf-love, Atimastillas flavicollis
- Spotted Greenbul, Ixonotus guttatus
- Swamp Palm Bulbul, Thescelocichla leucopleura
- Honeyguide Greenbul, Baeopogon indicator
- Sjostedt's Greenbul, Baeopogon clamans
- Joyful Greenbul, Chlorocichla laetissima
- Prigogine's Greenbul, Chlorocichla prigoginei
- Yellow-bellied Greenbul, Chlorocichla flaviventris
- Falkenstein's Greenbul, Chlorocichla falkensteini
- Simple Greenbul, Chlorocichla simplex
- Shelley's Greenbul, Arizelocichla masukuensis
- Kakamega Greenbul, Arizelocichla kakamegae
- Cameroon Greenbul, Arizelocichla montana
- Western Greenbul, Arizelocichla tephrolaema
- Mountain Greenbul, Arizelocichla nigriceps
- Uluguru Greenbul, Arizelocichla neumanni
- Black-browed Greenbul, Arizelocichla fusciceps
- Yellow-throated Greenbul, Arizelocichla chlorigula
- Olive-breasted Greenbul, Arizelocichla kikuyuensis
- Stripe-cheeked Greenbul, Arizelocichla milanjensis
- Olive-headed Greenbul, Arizelocichla olivaceiceps
- Stripe-faced Greenbul, Arizelocichla striifacies
Pycnonotinae: Bulbuls G.R. Gray, 1840
- Hairy-backed Bulbul, Tricholestes criniger
- Hook-billed Bulbul, Setornis criniger
- Yellow-bellied Bulbul, Alophoixus phaeocephalus
- Gray-cheeked Bulbul, Alophoixus bres
- Palawan Bulbul, Alophoixus frater
- Finsch's Bulbul, Alophoixus finschii
- White-throated Bulbul, Alophoixus flaveolus
- Puff-throated Bulbul, Alophoixus pallidus
- Ochraceous Bulbul, Alophoixus ochraceus
- Yellow-browed Bulbul, Acritillas indica
- Sulphur-bellied Bulbul, Iole palawanensis
- Buff-vented Bulbul, Iole crypta
- Charlotte's Bulbul, Iole charlottae
- Gray-eyed Bulbul, Iole propinqua
- Cachar Bulbul, Iole cacharensis
- Olive Bulbul, Iole viridescens
- Cinereous Bulbul, Hemixos cinereus
- Ashy Bulbul, Hemixos flavala
- Chestnut Bulbul, Hemixos castanonotus
- Nicobar Bulbul, Ixos nicobariensis
- Mountain Bulbul, Ixos mcclellandii
- Streaked Bulbul, Ixos malaccensis
- Sunda Bulbul, Ixos viridescens
- White-headed Bulbul, Cerasophila thompsoni
- Northern Golden-Bulbul, Thapsinillas longirostris
- Seram Golden-Bulbul, Thapsinillas affinis
- Buru Golden-Bulbul, Thapsinillas mysticalis
- Visayan Bulbul, Hypsipetes guimarasensis
- Zamboanga Bulbul, Hypsipetes rufigularis
- Yellowish Bulbul, Hypsipetes everetti
- Philippine Bulbul, Hypsipetes philippinus
- Mindoro Bulbul, Hypsipetes mindorensis
- Streak-breasted Bulbul, Hypsipetes siquijorensis
- Brown-eared Bulbul, Hypsipetes amaurotis
- Black Bulbul, Hypsipetes leucocephalus
- Square-tailed Bulbul, Hypsipetes ganeesa
- Moheli Bulbul, Hypsipetes moheliensis
- Seychelles Bulbul, Hypsipetes crassirostris
- Grand Comoro Bulbul, Hypsipetes parvirostris
- Reunion Bulbul, Hypsipetes borbonicus
- Malagasy Bulbul, Hypsipetes madagascariensis
- Mauritius Bulbul, Hypsipetes olivaceus
- Black-and-white Bulbul, Microtarsus melanoleucos
- Puff-backed Bulbul, Microtarsus eutilotus
- Gray-headed Bulbul, Microtarsus priocephalus
- Black-headed Bulbul, Microtarsus atriceps
- Andaman Bulbul, Microtarsus fuscoflavescens
- Blue-wattled Bulbul, Microtarsus nieuwenhuisii
- Yellow-wattled Bulbul, Microtarsus urostictus
- Crested Finchbill, Spizixos canifrons
- Collared Finchbill, Spizixos semitorques
- Black-crested Bulbul, Rubigula flaviventris
- Ruby-throated Bulbul, Rubigula dispar
- Flame-throated Bulbul, Rubigula gularis
- Black-capped Bulbul, Rubigula melanictera
- Bornean Bulbul, Rubigula montis
- Spectacled Bulbul, Rubigula erythropthalmos
- Scaly-breasted Bulbul, Rubigula squamata
- Gray-bellied Bulbul, Rubigula cyaniventris
- Stripe-throated Bulbul, Pycnonotus finlaysoni
- Straw-headed Bulbul, Pycnonotus zeylanicus
- Striated Bulbul, Pycnonotus striatus
- Cream-striped Bulbul, Pycnonotus leucogrammicus
- Spot-necked Bulbul, Pycnonotus tympanistrigus
- Olive-winged Bulbul, Pycnonotus plumosus
- Ashy-fronted Bulbul, Pycnonotus cinereifrons
- Streak-eared Bulbul, Pycnonotus blanfordi
- Cream-vented Bulbul, Pycnonotus simplex
- Asian Red-eyed Bulbul, Pycnonotus brunneus
- Flavescent Bulbul, Pycnonotus flavescens
- Yellow-vented Bulbul, Pycnonotus goiavier
- White-browed Bulbul, Pycnonotus luteolus
- Bare-faced Bulbul, Pycnonotus hualon
- Brown-breasted Bulbul, Pycnonotus xanthorrhous
- Light-vented Bulbul, Pycnonotus sinensis
- Red-whiskered Bulbul, Pycnonotus jocosus
- Styan's Bulbul, Pycnonotus taivanus
- Common Bulbul, Pycnonotus barbatus
- Somali Bulbul, Pycnonotus somaliensis
- Dodson's Bulbul, Pycnonotus dodsoni
- Dark-capped Bulbul, Pycnonotus tricolor
- African Red-eyed Bulbul, Pycnonotus nigricans
- Cape Bulbul, Pycnonotus capensis
- White-spectacled Bulbul, Pycnonotus xanthopygos
- White-eared Bulbul, Pycnonotus leucotis
- Himalayan Bulbul, Pycnonotus leucogenys
- Red-vented Bulbul, Pycnonotus cafer
- Sooty-headed Bulbul, Pycnonotus aurigaster
- Aceh Bulbul, Pycnonotus snouckaerti
- Orange-spotted Bulbul, Pycnonotus bimaculatus
- Yellow-throated Bulbul, Pycnonotus xantholaemus
- Yellow-eared Bulbul, Pycnonotus penicillatus