Australaves Ericson, 2012
This clade was one of the surprises found using DNA. It first popped up in Ericson et al. (2006). Since then, there have been numerous papers supporting it. There are the conventional DNA analyses of Ericson et al. (2006) and Hackett et al. (2008). Although these use the controversial 7th intron of the β-fibrogen gene, the clade also appears in Ericson et al.'s supplementary ESM-6 tree, which does not use the gene. Wang et al. (2012) use 30 nuclear introns (not including any introns of β-fibrogen). Their work also supports the notion that passerines and parrots are sisters, and that the falcons and seriemas are their closet relatives. Nabholz et al. (2011) also support the close relation between the passerines and parrots, although taxon sampling is very sparse and they do not consider the falcons. Passerimorphae also appear as a clade (under the name Eufalconimorphae) in Suh et al. (2011). They use the completely different approach of retroposon insertions to investigate avian phylogeny. They also note the similarities in the vocal apparatus and the fact that both parrots and many passeriformes learn their song. Finally, Jarvis et al. (2014) found it strongly supported in everything but the exon data (which don't support very much of anything).
CARIAMIFORMES Wagler, 1830
Cariamidae: Seriemas Bonaparte, 1850 (1836)
2 genera, 2 species HBW-3
The seriemas have sometimes been considered Gruiformes, but genetic evidence points elsewhere. Both Hackett et al. (2008) and Jarvis et al. (2014) found them sister to the remaining three orders, while Ericson et al. (2006a) suggest they belong in the Falconiformes. Either way, they are next in the list!
Although there are currently only two species of Seriemas, they are a remnant of a much larger group, the Cariamae. Like the Cariamidae, the Cariamae have traditionally been placed in the Gruiformes. Besides the Cariamidae, the Cariamae are thought to have included three now-extinct families: the Phorusrhacidae (S. America), Bathornithidae (N. America) and Idiornithidae (Europe). The oldest fossils are from Phorusrhacidae, dating back to the late Paleocene in Brazil, which matches well with the divergence estimate for the Cariamidae by Jarvis et al. (2014). The Phorusrhacidae are the well-known “terror birds” of ancient South America. See Alvarenga and Höfling (2003) for more on the Phorusrhacidae.
- Red-legged Seriema, Cariama cristata
- Black-legged Seriema, Chunga burmeisteri
FALCONIFORMES Sharpe, 1874
Falconidae: Falcons, Caracaras Leach, 1820
10 genera, 67 species HBW-2
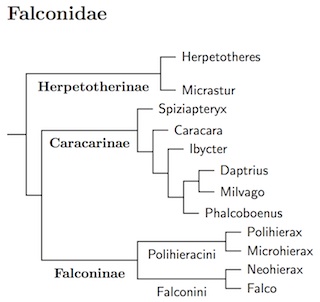 |
| Click for Falconidae tree |
|---|
The phylogeny is now based on the multigene analysis of Fuchs et al. (2015), who considered almost all of the species. Fuchs et al. (2015) calibrate their tree based on two fossils. They found that crown-group Falconidae date to the Oligocene. This is interesting as the crown-groups of their sister taxa (Psittaciformes plus Passeriformes) are older, much older. If the falcons are correctly placed as sister to the parrots and passeriformes, they must have originated in the early part of the Eocene or more likely Paleocene, perhaps 50-60 million years ago (some estimates make them even older). Yet only one lineage from the Oligocene (roughly 30 million years) is the ancestor of all the extant Falconidae, with all other Falconidae lineages from the previous 20-30 million years becoming extinct. Interestingly enough, the Falconiform fossil record is also quite sparse.
Previously, I'd used the papers by Griffiths (1999) and Griffiths et al. (2004) for the basic taxonomy of the Falconidae. This had been supplemented by Fuchs et al. (2011b) for the forest-falcon clade (Herpetotherinae), and Fuchs et al. (2012a) for the caracaras (Caracarinae).
Fuchs et al. (2015) found that many of the caracaras are closely related, with their common ancestor estimated to live less than 5 million years ago. The differences between them are not large, so I've merged the genera Milvago, and Phalcoboenus into Daptrius. Ibycter is somewhat different, not just because it is much less closely related to the Daptrius than the Daptrius are to one another, but in other ways. E.g., the syrinx is different from that of the Caracaras in the expanded Daptrius. See also SACC proposal #1038.
Fuchs et al. (2015) found that the two species of Polihierax are not closely related, with the Pygmy Falcon closer to Microhierax and the White-rumped Falcon nearer Falco. Accordingly, the White-rumped Falcon is placed in the monotypic genus Neohierax (Swann, 1922). Falco itself seems to have originated much more recently, with a common ancestor most likely between 5 and 8 million years ago, a good reason to leave all of Falco in a single genus. In contrast, the Neohierax/Falco split was likely 10-15 million years ago. To further emphasize the division in Falconinae, I've recognized two tribes within it: Polihieracini and Falconini.
The African kestrel clade was studied by Groombridge et al. (2002). Based on their work, Rock Kestrel must be split from Eurasian Kestrel because the Australian Kestrel is split. The Hierofalco complex was examined in detail by Wink et al. (2004a). In spite of sampling many individuals, they were unable to produce a clean phylogeny. Although this may mean some of the these species should be lumped, they believe the confused situation is due to ancient hybridization. Further analysis is required to sort out the Hierofalco complex. This remains true even after Fuchs et al.'s (2015) analysis. The order of Falco itself is now based on Fuchs et al. (2015) rather than Wink and Sauer-Gürth (2004).
Herpetotherinae: Forest-Falcons Lesson, 1843
- Laughing Falcon, Herpetotheres cachinnans
- Buckley's Forest-Falcon, Micrastur buckleyi
- Collared Forest-Falcon, Micrastur semitorquatus
- Slaty-backed Forest-Falcon, Micrastur mirandollei
- Barred Forest-Falcon, Micrastur ruficollis
- Cryptic Forest-Falcon, Micrastur mintoni
- Plumbeous Forest-Falcon, Micrastur plumbeus
- Lined Forest-Falcon, Micrastur gilvicollis
Caracarinae: Caracaras d'Orbigny, 1837
- Spot-winged Falconet, Spiziapteryx circumcincta
- Southern Caracara / Southern Crested Caracara, Caracara plancus
- Crested Caracara / Northern Crested Caracara, Caracara cheriway
- †Guadalupe Caracara, Caracara lutosa
- Red-throated Caracara, Ibycter americanus
- Black Caracara, Daptrius ater
- Yellow-headed Caracara, Daptrius chimachima
- Chimango Caracara, Daptrius chimango
- Striated Caracara, Daptrius australis
- Carunculated Caracara, Daptrius carunculatus
- Mountain Caracara, Daptrius megalopterus
- White-throated Caracara, Daptrius albogularis
Falconinae: Falcons Leach, 1820
Polihieracini Peters, 1931
- Pygmy Falcon, Polihierax semitorquatus
- Philippine Falconet, Microhierax erythrogenys
- Collared Falconet, Microhierax caerulescens
- Pied Falconet, Microhierax melanoleucos
- Black-thighed Falconet, Microhierax fringillarius
- White-fronted Falconet, Microhierax latifrons
Falconini Leach, 1820
- White-rumped Falcon, Neohierax insignis
- Banded Kestrel, Falco zoniventris
- Lesser Kestrel, Falco naumanni
- Fox Kestrel, Falco alopex
- Greater Kestrel, Falco rupicoloides
- Seychelles Kestrel, Falco araeus
- Malagasy Kestrel, Falco newtoni
- Reunion Kestrel, Falco duboisi
- Mauritius Kestrel, Falco punctatus
- Rock Kestrel, Falco rupicolus
- Spotted Kestrel, Falco moluccensis
- Eurasian Kestrel / Common Kestrel, Falco tinnunculus
- Nankeen Kestrel, Falco cenchroides
- American Kestrel, Falco sparverius
- Red-footed Falcon, Falco vespertinus
- Amur Falcon, Falco amurensis
- Merlin, Falco columbarius
- Gray Kestrel, Falco ardosiaceus
- Dickinson's Kestrel, Falco dickinsoni
- Brown Falcon, Falco berigora
- Bat Falcon, Falco rufigularis
- Orange-breasted Falcon, Falco deiroleucus
- Australian Hobby, Falco longipennis
- Oriental Hobby, Falco severus
- Eleonora's Falcon, Falco eleonorae
- Sooty Falcon, Falco concolor
- Eurasian Hobby, Falco subbuteo
- African Hobby, Falco cuvierii
- New Zealand Falcon, Falco novaeseelandiae
- Aplomado Falcon, Falco femoralis
- Red-necked Falcon, Falco chicquera
- Prairie Falcon, Falco mexicanus
- Gray Falcon, Falco hypoleucos
- Taita Falcon, Falco fasciinucha
- Peregrine Falcon, Falco peregrinus
- Barbary Falcon, Falco pelegrinoides
- Lanner Falcon, Falco biarmicus
- Laggar Falcon, Falco jugger
- Black Falcon, Falco subniger
- Gyrfalcon, Falco rusticolus
- Saker Falcon, Falco cherrug
PSITTACIFORMES Wagler, 1830
Ericson et al. (2006a), Hackett et al. (2008), and Jarvis et al. (2014) all identified the parrots as the sister group to the passeriformes.
The overall structure of the Psittaciformes follows Schirtzinger et al. (2012), Schweitzer et al. (2010), and Wright et al. (2008). Christidis and Boles (2008) contains detailed information on Australian parrots. Brown and Toft (1999) and White et al. (2011) provided further information concerning the Cockatoos; Groombridge et al. (2004) and Kundu et al. (2012) were helpful with the Psittaculini; the placement of the Ground and Night Parrots is inspired by Leeton et al. (1994). Tavares et al. (2006) gives a nice overview of the Arini, with Kirchman et al. (2012) providing additional detail. The papers by Ribas et al. (2005, 2007a, b) were helpful concerning the basal Androglossini, while Russello and Amato (2004) was consulted concerning the Amazona parrots. Russello and Amato found that the for last four (possibly five) Amazona species, the genetic data does not seem to match existing subspecies.
Although crown group parrots appear to originate in the Australo-Pacific region, there is plenty of fossil evidence that the early parrots were much more widespread. Stem parrot fossils have been found from the early Eocene of India (Vastanavis), Europe (Halcyornithidae, Psittacopes), and North America (Tynskya), see Mayr (2009), and Mayr et al., (2013).
The extant order is summed up in the following tree diagram.
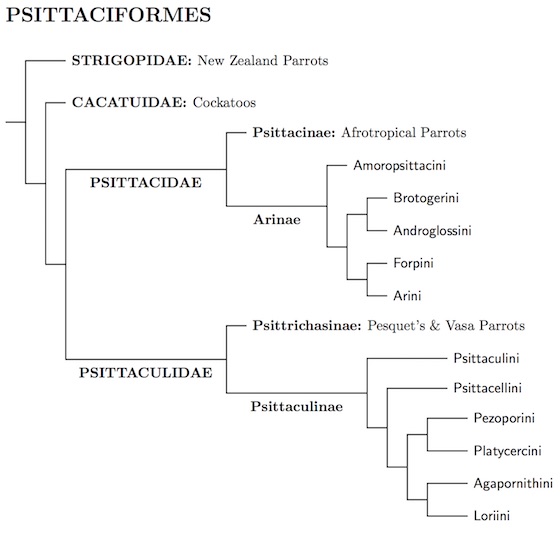 |
| Click for genus-level tree for Psittaciformes |
|---|
Strigopidae: New Zealand Parrots Bonaparte, 1849
2 genera, 4 species Not HBW Family
I had earlier used the name Nestoridae, which seemed to be the most common usage for a family containing both Strigops and Nestor. However, both family-group names were introduced at the same time (Bonaparte, 1849), and they have normally been kept separated enough that the issue of priority doesn't arise. I don't have access to Bonaparte (1849), but Bock (1994) does not mention priority between them, listing them as Strigopinae and Nestorinae. Christidis and Boles (2008) also state they have equal priority. Assuming that is correct, the determination of priority falls to the first reviser. That's Bonaparte himself the following year, when he gave priority to Strigopidae by listing Nestorinae as a subfamily (1850, Vol. 1, p. 8).
- Kakapo, Strigops habroptila
- Kea, Nestor notabilis
- Norfolk Kaka, Nestor productus
- New Zealand Kaka, Nestor meridionalis
Cacatuidae: Cockatoos G.R. Gray, 1840 (1825)
7 genera, 21 species HBW-4
The order within the cockatoos is based on White et al. (2011), with some help from Brown and Toft (1999). White et al. (2011) found that the Cockatiel was basal, and I follow their multigene analysis (as well as traditional thinking). Note however that Brown and Toft found a different position for the Cockatiel, which also received limited support from Wright et al. (2008).
Three of the black-cockatoos have been moved to the genus Zanda (Mathews 1913, type bauinii) due to substantial separation between them and the Calyptorhynchus black-cockatoos. White et al. (2011) estimate the most recent common ancestor at about 15 million years ago. Interesting, the current Zanda diversity appeared very recently, whereas the split between the two Calyptorhynchus is of much longer standing.
- Cockatiel, Nymphicus hollandicus
- Red-tailed Black-Cockatoo, Calyptorhynchus banksii
- Glossy Black-Cockatoo, Calyptorhynchus lathami
- Yellow-tailed Black-Cockatoo, Zanda funerea
- Carnaby's Black-Cockatoo / Short-billed Black-Cockatoo, Zanda latirostris
- Baudin's Black-Cockatoo / Long-billed Black-Cockatoo, Zanda baudinii
- Palm Cockatoo, Probosciger aterrimus
- Gang-gang Cockatoo, Callocephalon fimbriatum
- Galah, Eolophus roseicapilla
- Major Mitchell's Cockatoo, Cacatua leadbeateri
- Yellow-crested Cockatoo, Cacatua sulphurea
- Sulphur-crested Cockatoo, Cacatua galerita
- Blue-eyed Cockatoo, Cacatua ophthalmica
- Salmon-crested Cockatoo, Cacatua moluccensis
- White Cockatoo, Cacatua alba
- Red-vented Cockatoo, Cacatua haematuropygia
- Tanimbar Corella, Cacatua goffiniana
- Solomons Cockatoo, Cacatua ducorpsii
- Long-billed Corella, Cacatua tenuirostris
- Western Corella, Cacatua pastinator
- Little Corella, Cacatua sanguinea
Psittacidae: African and American Parrots Rafinesque, 1815
37 genera, 174 species HBW-4
Schirtzinger et al. (2012), Schweizer et al. (2010) and Wright et al. (2008) identify 5 deep clades in the the remaining parrots: the vasa-parrots, Pesquet's Parrot, an Old World/Australasian clade (Psittaculinae), an African clade (Psittacinae), and an American clade (Arinae). Pesquet's Parrot and the vasa parrots seem to be each other's closet relatives, and we unite them in the subfamily Psittrichasinae. The appear to be sister groups, as do the Psittrichasinae and Psittaculinae, leaving us with two major clades: Psittacidae (Psittacinae and Arinae) and Psittaculidae (Psittrichasinae and Psittaculinae).
How deep are the five clades? Wright et al. (2008) gave two different timelines. In one, they date from the late Cretaceous to the early Paleocene, roughly 65 million years ago. They also provide a later dating, in the Eocene, about 40 million years ago. The earlier dating suppose that the New Zealand Parrots date from the separation of New Zealand from Gondwana, over 80 million years ago. The later dating puts the split of the New Zealand Parrots about 50 million years ago, in the Eocene. Joseph et al. (2012), who seem to prefer an early dating, recommend ranking Psittacinae, Psittrichasinae, and Psittaculinae as families rather than subfamiles. Personally, I prefer the later dating as it fits better with the fossil record. It also fits better with Jarvis et al. (2014), who estimate the parrot/perching bird split at about 55 mya. I follow Howard and Moore 4th ed. and SACC rather than Joseph et al., and use only two families: Psittacidae and Psittaculidae.
Of the two main clades, Psittaculidae is slightly larger, so we put it last in the linear order.
Psittacinae: Afrotropical Parrots Rafinesque, 1815
- Gray Parrot, Psittacus erithacus
- Brown-necked Parrot, Poicephalus fuscicollis
- Cape Parrot, Poicephalus robustus
- Red-fronted Parrot, Poicephalus gulielmi
- Meyer's Parrot, Poicephalus meyeri
- Rueppell's Parrot, Poicephalus rueppellii
- Brown-headed Parrot, Poicephalus cryptoxanthus
- Niam-niam Parrot, Poicephalus crassus
- Red-bellied Parrot, Poicephalus rufiventris
- Senegal Parrot, Poicephalus senegalus
- Yellow-fronted Parrot, Poicephalus flavifrons
Arinae: Neotropical Parrots G.R. Gray, 1840 (1825)
The Neotropical parrots, Arinae, fall into 5 subclades: Amoropsittacini, Brotogerini, the clade of large short-tailed parrots, Androglossini, Forpus parrotlets (Forpini), and a narrowly circumscribed conure/macaw clade Arini. The relationships between them are not fully understood at this time—the various relevant papers give conflicting topologies (de Kloet and de Kloet, 2005; Tavares et al., 2006; Wright et al., 2008, Schirtzinger et al., 2012). However, the Tavares et al. (2006) topology seems to fit best with the other papers, and I have adopted it.
In the Tavares et al. topology, Amoropsittacini is the basal group. Schirtzinger et al. (2012) also included Psilopsiagon in their analysis, putting it here in the Amoropsittacini. The Brotogerini appear to be affiliated with the Androglossini, and the Forpini with the core Arini.
Amoropsittacini Brereton, 1963
- Gray-hooded Parakeet, Psilopsiagon aymara
- Mountain Parakeet, Psilopsiagon aurifrons
- Tepui Parrotlet, Nannopsittaca panychlora
- Amazonian Parrotlet / Manu Parrotlet, Nannopsittaca dachilleae
- Barred Parakeet, Bolborhynchus lineola
- Rufous-fronted Parakeet, Bolborhynchus ferrugineifrons
- Andean Parakeet, Bolborhynchus orbygnesius
- Lilac-tailed Parrotlet, Touit batavicus
- Scarlet-shouldered Parrotlet, Touit huetii
- Red-fronted Parrotlet, Touit costaricensis
- Blue-fronted Parrotlet, Touit dilectissimus
- Sapphire-rumped Parrotlet, Touit purpuratus
- Brown-backed Parrotlet, Touit melanonotus
- Golden-tailed Parrotlet, Touit surdus
- Spot-winged Parrotlet, Touit stictopterus
Brotogerini Wolters, 1975
The ordering of the Brotegerini is based on Ribas et al. (2009). They found that the Brotogeris parakeets fall into two groups of 4 species each. Not surprisingly, the genetic distance between chiriri and versicolurus was fairly small, with the amount being consistent with treatment either as separate species or subspecies of a single species.
- Monk Parakeet, Myiopsitta monachus
- Plain Parakeet, Brotogeris tirica
- Tui Parakeet, Brotogeris sanctithomae
- White-winged Parakeet / Canary-winged Parakeet, Brotogeris versicolurus
- Yellow-chevroned Parakeet, Brotogeris chiriri
- Gray-cheeked Parakeet, Brotogeris pyrrhoptera
- Orange-chinned Parakeet, Brotogeris jugularis
- Cobalt-winged Parakeet, Brotogeris cyanoptera
- Golden-winged Parakeet, Brotogeris chrysoptera
Androglossini Sundevall, 1872
The series of papers by Ribas et al. (2005, 2007a, b) and Russello and Amato (2004) have done much to clarify the situation in the Androglossini. The parrots from Pionopsitta to Pyrilia were studied by Ribas et al. (2005). The ordering of the Pionus parrots reflects the results of Ribas et al. (2007a). The results also support a possible split of White-capped Parrot, Pionus seniloides. The Hapalopsittaca order is based on Quintero et al. (2013).
Wenner et al. (2012) found a substantial amount of genetic structure in the Mealy Amazon, Amazona farinosa. The Central American populations to west Panama (guatemalae and virenticeps) formed a separate clade from those in the Darien and South America (inornata and farinosa/chapmani), with 3.5-5.4% genetic distance in cytochrome-b. Such differences usually indicate separate species, and I treat them as such: the Northern Mealy Amazon, Amazona guatemalae, and Southern Mealy Amazon, Amazona farinosa. Some uncertainty remains here as they did not look in detail at the situation within Panama where the two clades are in contact. There's a slight plumage difference, but it doesn't seem strong enough to provide a separating mechanism. It is possible that they are one biological species in spite of the DNA differences. Also, contrary to its treatment in recent checklists, the DNA of the Choco-Darien form inornata was clearly distinct from that of farinosa/chapmani and should at least be recognized as a subspecies.
Yellow-headed Amazons: The big issue here is the Yellow-headed Amazon complex—the last eight species in Androglossini. This group has long been controversial, and genetic studies have revealed new complexities. The papers by Rusello et al. (2004) and Eberhard and Bermingham (2004) showed the Turquoise-fronted Amazon is a member of this complex. Ribas et al. (2007b) sampled many more of the South American birds in an effort to clear up the mysteries of this group, but the relationships there remain confusing. Caparroz et al. (2009) sampled some additional locations. Finally, Urantówka et al. (2014) helped clarify matters concerning A. barbadensis.
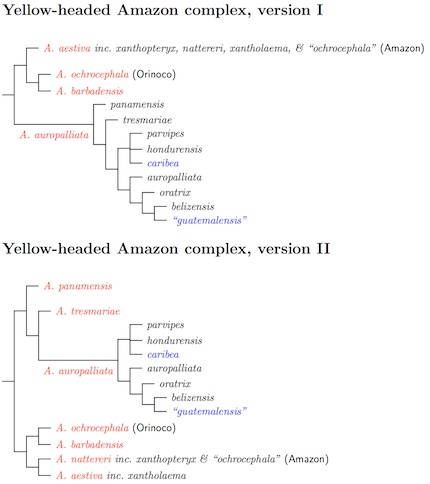 There is substantial variation in the Yellow-headed Amazon complex.
The species of the complex are divided into 15 recognized subspecies and one
informal subspecies. This does not capture all of the variation as
some very distinct parrots, such as the Auás of Bahia, Brazil are
not recognized as subspecies. The Auá is considered part of
nominate Turquoise-fronted aestiva in spite of its extensive
yellow (or orange!) bib. Monroe and Howell (1966) surveyed the variation
in Middle American members of the Yellow-headed complex.
There is substantial variation in the Yellow-headed Amazon complex.
The species of the complex are divided into 15 recognized subspecies and one
informal subspecies. This does not capture all of the variation as
some very distinct parrots, such as the Auás of Bahia, Brazil are
not recognized as subspecies. The Auá is considered part of
nominate Turquoise-fronted aestiva in spite of its extensive
yellow (or orange!) bib. Monroe and Howell (1966) surveyed the variation
in Middle American members of the Yellow-headed complex.
- aestiva
- auropalliata
- barbadensis inc. rothschildi
- belizensis
- caribaea
- hondurensis
- magna
- nattereri
- ochrocephala
- oratrix
- panamensis
- parvipes
- tresmariae
- xantholaema
- xanthopteryx
- “guatemalensis” (not formally described)
Various divisions into species have been used. Needless to say, the DNA has had some surprises for us. The two figures show two arrangements that are consistent with current DNA evidence. You'll note that ochrocephala appears twice. Although sampling has not been detailed enough to draw the lines sharply, it seems that the birds in the Orinoco basin are different from those in the Amazon. The type of ochrocephala comes from Colombia, in the Orinoco drainage. I can't find an alternate name for the Amazonian ochrocephala, so I've labelled them as ochrocephala (Amazon) and ochrocephala (Orinoco).
The literature refers to a Central (meaning Middle) American clade, Northern South American (NSA) clade and two South American clades (SA1 and SA2). In version II of the diagram, the first 3 species in the Middle American clade (note that the range of panamensis extends into Colombia, north and west of the Andes), the next two are the NSA clade, and the last two are the South American clades (SA2 and SA1, respectively).
The Middle American subspecies currently in oratrix and auropalliata are fairly closely related, and could be reasonably grouped under A. auropalliata (auropalliata Lesson, 1842 has priority over oratrix Ridgway, 1887). That gives us four species as shown in version one (the species names are in red). Some have argued that the Tres Marias Amazon should be considered a separate species, and version II shows how such a tree would look. Although parvipes, hondurensis, and caribea are on a different branch than either auropalliata or oratrix, support for this seems fairly weak. Urantówka et al. (2014) has a different topology in Figure 4 where all three group with auropalliata. Until more data is available, I continue to recognize the two traditional species, auropalliata (Yellow-naped) and oratrix (Yellow-headed) for the auropalliata group. However, the Tres Marias Amazon, Amazona tresmariae, appears to be more basal than any of the auropalliata/oratrix group and is now considered a distinct species.
Although panamensis is generally considered part of South American ochrocephala, it is the basal member of the Middle American clade. Accordingly, I have elevated panamensis to species status as the Panama Amazon.
This brings us to the South American clades. These clades involve not just the traditional Yellow-headed complex but also the Turquoise-fronted Amazon, Amazona aestiva. The clades do not map well into traditional taxonomies.
The NSA clade includes the Yellow-shouldered Amazon, Amazona barbadensis, and ochrocephala that have been sampled from Colombia (east of the Andes), Venezuela, and Guyana. The thing that unites this birds is that all are from the Orinoco basin. Since the type of ochrocephala is from the Orinoco drainage in Colombia, the ochrocephala in the NSA clade are the true ochrocephala. There seems to be over 2% DNA difference between the NSA clade and the SA clades. The true Amazona ochrocephala is separated from other “ochrocephala” as the Orinocan Yellow-crowned Amazon.
The two South America clades are trickier. The genetic distance is smaller (about 1%), but there's a lot of variation in plumage (it could even be called excessive) and these have been traditionally considered two species. But the DNA simply does not match traditional species divisions. It could be that the DNA is wrong. This is based on mitochondrial DNA, and very little nuclear DNA has been considered. Eberhard and Bermingham (2004) did use a few samples of nuclear DNA. It seems to show a different topology something other than SA1 and SA2. Eberhard and Bermingham performed a Shimodaira-Hasegawa test and it failed to reject the mitochondrial topology. Additional nuclear data could lead to a different result, but for now, the mitochondrial topology looks good.
The SA2 clade consists of Amazonian “ochrocephala”, nattereri, and surprisingly xanthopteryx (usually grouped in Turquoise-fronted Amazon). The sampling of “ochrocephala” from the Amazon is sparse, including 4 birds from Macapá (north side of the Amazon), and 2 from Altimira on the Xingú (south of the Amazon). There doesn't appear to be an alternate name for “ochrocephala”, so nattereri has priority. Together, these form the Amazonian Yellow-crowned Amazon, Amazona nattereri.
The SA1 clade includes nominate aestiva and xantholaema (usually grouped with ochrocephala). Although restructured, it's still Turquoise-fronted Amazon.
- Red-capped Parrot / Pileated Parrot, Pionopsitta pileata
Click for Androglossini tree - Blue-bellied Parrot, Triclaria malachitacea
- Black-winged Parrot, Hapalopsittaca melanotis
- Rusty-faced Parrot, Hapalopsittaca amazonina
- Indigo-winged Parrot / Fuertes's Parrot, Hapalopsittaca fuertesi
- Red-faced Parrot, Hapalopsittaca pyrrhops
- Brown-hooded Parrot, Pyrilia haematotis
- Rose-faced Parrot, Pyrilia pulchra
- Saffron-headed Parrot, Pyrilia pyrilia
- Orange-cheeked Parrot, Pyrilia barrabandi
- Caica Parrot, Pyrilia caica
- Bald Parrot, Pyrilia aurantiocephala
- Vulturine Parrot, Pyrilia vulturina
- Short-tailed Parrot, Graydidascalus brachyurus
- Yellow-faced Parrot, Alipiopsitta xanthops
- Dusky Parrot, Pionus fuscus
- Red-billed Parrot, Pionus sordidus
- Scaly-headed Parrot, Pionus maximiliani
- White-capped Parrot, Pionus seniloides
- Plum-crowned Parrot, Pionus tumultuosus
- Blue-headed Parrot, Pionus menstruus
- White-crowned Parrot, Pionus senilis
- Bronze-winged Parrot, Pionus chalcopterus
- White-fronted Parrot / White-fronted Amazon, Amazona albifrons
- Yellow-lored Parrot / Yucatan Amazon, Amazona xantholora
- Black-billed Parrot / Black-billed Amazon, Amazona agilis
- Yellow-billed Parrot / Yellow-billed Amazon, Amazona collaria
- Hispaniolan Parrot / Hispaniolan Amazon, Amazona ventralis
- Cuban Parrot / Cuban Amazon, Amazona leucocephala
- Puerto Rican Parrot / Puerto Rican Amazon, Amazona vittata
- Festive Parrot / Festive Amazon, Amazona festiva
- Vinaceous-breasted Parrot / Vinaceous-breasted Amazon, Amazona vinacea
- Tucuman Parrot / Tucuman Amazon, Amazona tucumana
- Red-spectacled Parrot / Red-spectacled Amazon, Amazona pretrei
- Imperial Parrot / Imperial Amazon, Amazona imperialis
- Red-tailed Parrot / Red-tailed Amazon, Amazona brasiliensis
- Orange-winged Parrot / Orange-winged Amazon, Amazona amazonica
- St. Vincent Parrot / St. Vincent Amazon, Amazona guildingii
- Red-crowned Parrot / Red-crowned Amazon, Amazona viridigenalis
- Lilac-crowned Parrot / Lilac-crowned Amazon, Amazona finschi
- Red-lored Parrot / Red-lored Amazon, Amazona autumnalis
- Scaly-naped Parrot / Scaly-naped Amazon, Amazona mercenarius
- Kawall's Parrot / Kawall's Amazon, Amazona kawalli
- Northern Mealy Parrot / Northern Mealy Amazon, Amazona guatemalae
- Southern Mealy Parrot / Southern Mealy Amazon, Amazona farinosa
- Blue-cheeked Parrot / Blue-cheeked Amazon, Amazona dufresniana
- Red-browed Parrot / Red-browed Amazon, Amazona rhodocorytha
- Red-necked Parrot / Red-necked Amazon, Amazona arausiaca
- St. Lucia Parrot / St. Lucia Amazon, Amazona versicolor
- Panama Parrot / Panama Amazon, Amazona panamensis
- Tres Marias Parrot / Tres Marias Amazon, Amazona tresmariae
- Yellow-headed Parrot / Yellow-headed Amazon, Amazona oratrix
- Yellow-naped Parrot / Yellow-naped Amazon, Amazona auropalliata
- Orinocan Yellow-crowned Parrot / Orinocan Yellow-crowned Amazon, Amazona ochrocephala
- Yellow-shouldered Parrot / Yellow-shouldered Amazon, Amazona barbadensis
- Amazonian Yellow-crowned Parrot / Amazonian Yellow-crowned Amazon, Amazona nattereri
- Turquoise-fronted Parrot / Turquoise-fronted Amazon, Amazona aestiva
Forpini: Forpus Parrotlets Brereton, 1963
The arrangement within Forpus is based on B.T. Smith et al. (2013). The species limits may need some adjustment. In particular, the Blue-winged Parrotlet, Forpus xanthopterygius, may not be monophyletic.
- Dusky-billed Parrotlet, Forpus modestus
- Blue-winged Parrotlet, Forpus xanthopterygius
- Mexican Parrotlet, Forpus cyanopygius
- Green-rumped Parrotlet, Forpus passerinus
- Spectacled Parrotlet, Forpus conspicillatus
- Pacific Parrotlet, Forpus coelestis
- Yellow-faced Parrotlet, Forpus xanthops
Arini: Conures and Macaws G.R. Gray, 1840 (1825)
Tavares et al. (2006) found strong support for a basal Deroptyus-Pionites grouping, sister to the remaining Arini. This arrangement also got mixed support from Wright et al. (2008), and is adopted here.
The Yellow-eared Parrot, Ognorhynchus icterotis, doesn't seem to have been included in any genetic studies. It probably groups with the remaining Arini, but it's true position is unresolved.
Rhynchopsitta may group with Pyrrhura (Tavares et al., 2006), or maybe not (Kirchman et al., 2012). The arrangement of the Pyrrhura parakeets is based on Ribas et al. (2006).
The southern genera Enicognathus, Cyanoliseus, and Anodorhynchus seem to branch successively, bringing us to the macaws, Aratinga parakeets, and allies. This group includes a well-supported macaw clade, Cyanopsitta though Ara, and a small, well-supported grouping of Leptosittaca, Diopsittaca, and Guaruba. Urantowka et al. (2013) found that the Blue-crowned Parakeet is sister to Diopsittaca. I've placed it in the monotypic genus Thectocercus (Ridgway 1912).
Aratinga: The traditional version of Aratinga is not a monophyletic group (Ribas and Miyaki, 2004). The problem is how to separate them, and how they fit with the other two groups. Recently, Kirchman et al. (2012) included enough species in their analysis to split up Aratinga in a reasonable way. Support for the overall phylogeny, as followed here, is mediocre. I think the division of Aratinga is on more solid ground.
Kirchman et al. show three clades containing Aratinga parakeets. One clade is sister to the Carolina Parakeet, Conuropsis carolinensis. It includes the type species of Aratinga, the Sun Parakeet, Aratinga solstitialis and also auricapillus. Ribas and Miyaki (2004) showed that jandaya is sister to auricapillus, so it belongs here too, as does the the recently split maculata (see Silveira et al., 2005. Note that the scientific name has been corrected to A. maculata from A. pintoi by Nemésio and Rasmussen, 2009). Kirchman et al. show the Nanday Parakeet (usually Nandayus nenday) as the basal member of the group. However, a look at Ribas and Miyaki suggests that the Dusky-headed Parakeet, Aratinga weddellii, is the true basal member. Accordingly, I've merged Nandayus into Aratinga, which applies to these dark-billed parakeets. The whole lot may be sister to the macaw clade.
The horn-billed, green-winged, generally red and green “Aratinga” also appear to form a clade. The oldest available genus name is Psittacara (Vigors 1825, type leucophthalmus). Kirchman et al. (2012) found the Psittacara sister to the Leptosittaca-Guaruba group.
Based on Olson (2015), the extinct Puerto Rican Parakeet, Psittacara maugei, has been split from Hispaniolan Parakeet, Psittacara chloropterus.
The other clade of former Aratinga includes aurea, nana, and pertinax. Comparison with Ribas and Miyaki suggests that cactorum belongs here too. Since canicularis is thought to form a superspecies with aurea, it also goes in this group. The oldest available name seems to be Eupsittula (Bonaparte 1853, type canicularis). Kirchman et al. show this clade as sister to Rhynchopsitta. Maybe, but stronger evidence from other papers (e.g., Tavares et al., 2006) put Rhynchopsitta elsewhere, and so do I.
- Red-fan Parrot, Deroptyus accipitrinus
- Black-headed Parrot, Pionites melanocephalus
- White-bellied Parrot, Pionites leucogaster
- Yellow-eared Parrot, Ognorhynchus icterotis
- Thick-billed Parrot, Rhynchopsitta pachyrhyncha
- Maroon-fronted Parrot, Rhynchopsitta terrisi
- Blue-throated Parakeet / Ochre-marked Parakeet, Pyrrhura cruentata
- Blaze-winged Parakeet, Pyrrhura devillei
- Maroon-bellied Parakeet, Pyrrhura frontalis
- Pearly Parakeet, Pyrrhura lepida
- Crimson-bellied Parakeet, Pyrrhura perlata
- Green-cheeked Parakeet, Pyrrhura molinae
- Pfrimer's Parakeet, Pyrrhura pfrimeri
- Gray-breasted Parakeet, Pyrrhura griseipectus
- Maroon-faced Parakeet / White-eared Parakeet, Pyrrhura leucotis
- Painted Parakeet, Pyrrhura picta
- Santarem Parakeet, Pyrrhura amazonum
- Bonaparte's Parakeet, Pyrrhura lucianii
- Rose-fronted Parakeet, Pyrrhura roseifrons
- Santa Marta Parakeet, Pyrrhura viridicata
- Fiery-shouldered Parakeet, Pyrrhura egregia
- Maroon-tailed Parakeet, Pyrrhura melanura
- El Oro Parakeet, Pyrrhura orcesi
- Black-capped Parakeet, Pyrrhura rupicola
- White-necked Parakeet / White-breasted Parakeet, Pyrrhura albipectus
- Brown-breasted Parakeet / Flame-winged Parakeet, Pyrrhura calliptera
- Ochre-marked Parakeet / Blood-eared Parakeet, Pyrrhura hoematotis
- Rose-headed Parakeet / Rose-crowned Parakeet, Pyrrhura rhodocephala
- Sulphur-winged Parakeet, Pyrrhura hoffmanni
- Austral Parakeet, Enicognathus ferrugineus
- Slender-billed Parakeet, Enicognathus leptorhynchus
- Burrowing Parakeet / Burrowing Parrot, Cyanoliseus patagonus
- Hyacinth Macaw, Anodorhynchus hyacinthinus
- Glaucous Macaw, Anodorhynchus glaucus
- Indigo Macaw / Lear's Macaw, Anodorhynchus leari
- Brown-throated Parakeet, Eupsittula pertinax
- Cactus Parakeet / Caatinga Parakeet, Eupsittula cactorum
- Olive-throated Parakeet, Eupsittula nana
- Orange-fronted Parakeet, Eupsittula canicularis
- Peach-fronted Parakeet, Eupsittula aurea
- Golden-plumed Parakeet, Leptosittaca branickii
- Golden Parakeet, Guaruba guarouba
- Blue-crowned Parakeet, Thectocercus acuticaudatus
- Red-shouldered Macaw, Diopsittaca nobilis
- White-eyed Parakeet, Psittacara leucophthalmus
- Scarlet-fronted Parakeet, Psittacara wagleri
- Mitred Parakeet, Psittacara mitratus
- Red-masked Parakeet, Psittacara erythrogenys
- Crimson-fronted Parakeet / Finsch's Parakeet, Psittacara finschi
- Green Parakeet, Psittacara holochlorus
- Pacific Parakeet, Psittacara strenuus
- Cuban Parakeet, Psittacara euops
- Hispaniolan Parakeet, Psittacara chloropterus
- Puerto Rican Parakeet, Psittacara maugei
- Carolina Parakeet, Conuropsis carolinensis
- Dusky-headed Parakeet, Aratinga weddellii
- Nanday Parakeet, Aratinga nenday
- Sun Parakeet, Aratinga solstitialis
- Sulphur-breasted Parakeet, Aratinga maculata
- Jandaya Parakeet, Aratinga jandaya
- Golden-capped Parakeet, Aratinga auricapillus
- Spix's Macaw, Cyanopsitta spixii
- Red-bellied Macaw, Orthopsittaca manilatus
- Blue-winged Macaw, Primolius maracana
- Blue-headed Macaw, Primolius couloni
- Yellow-collared Macaw / Golden-collared Macaw, Primolius auricollis
- Cuban Macaw, Ara tricolor
- Blue-and-yellow Macaw, Ara ararauna
- Blue-throated Macaw, Ara glaucogularis
- Military Macaw, Ara militaris
- Great Green Macaw, Ara ambiguus
- Scarlet Macaw, Ara macao
- Red-and-green Macaw, Ara chloropterus
- Red-fronted Macaw, Ara rubrogenys
- Chestnut-fronted Macaw, Ara severus
Psittaculidae: Old World Parrots Vigors, 1825
49 genera, 192 species HBW-4
Psittaculidae contains two subfamilies: Psittrichasinae and Psittaculinae.
Psittrichasinae: Pesquet's & Vasa Parrots von Boetticher, 1959 (1854)
Kundu et al. (2012) examined a variety of Indian Ocean parrots. They found that that extinct Mascarene Parrot, Mascarinus mascarinus, belongs in the same genus as the vasa parrots. As Mascarinus (Lesson, 1830) is an older name that Coracopsis (Wagler, 1832), the entire group takes the genus name Mascarinus. The subfamily name remains unchanged. Kundu et al. also found that the Seychelles Black Parrot, usually considered a race of the Lesser Vasa Parrot, is fairly distant from the Lesser Vasa Parrot. Accordingly, I treat it as a separate species, Mascarinus barklyi.
- Pesquet's Parrot, Psittrichas fulgidus
- Greater Vasa-Parrot, Mascarinus vasa
- Seychelles Black Parrot, Mascarinus barklyi
- Mascarene Parrot, Mascarinus mascarinus
- Lesser Vasa-Parrot, Mascarinus niger
Psittaculinae Vigors, 1825
Why Six Tribes? Schweizer et al. (2010) and Wright et al. (2008) both found 5 major clades within the Psittaculinae, although neither group included Pezoporus or Psittacella in their analysis.
So where does Pezoporus go? Based on Leeton et al. (1994) and Miyaki et al. (1998), it seems to be sister to Neopsephotus + Neophema, and I treat all three as a single clade (Pezoporini). However, this is not as solid as it might be because neither included the Agapornithini in their analysis, which Wright et al. had as sister group to Neopsephotus + Neophema. More recently, Joseph et al. (2011) included all the relevant taxa in a multigene analysis and established that Pezoporus is sister to Neopsephotus + Neophema.
Joseph et al. (2011) also showed that the Psittacella tiger-parrots form a separate clade. This gives us 6 clades: Psittaculini; Psittacellini; Pezoporini; Platycercini; Agapornithini; Loriini.
Psittaculini: Pygmy-Parrots, King Parrots, Racket-tails, Ringnecks: The Psittaculini are basal to the others tribes in Psittaculinae. This tribe contains four main pieces, the pygmy parrots (Micropsitta), the king parrot group, Alisterus through Polytelis, the Prioniturus racket-tails, and the ringneck group, Eclectus through Psittacula, possibly excepting the extinct Lophopsittacus and Necropsittacus. These two, which might be each other's closest relatives have long been believed to be related to Psittacula. However, it is possible that they are closer to the Mascarene Parrot and Vasa Parrots.
There is some question about the monophyly of Polytelis, although Joseph et al. (2011) and Schweizer et al. (2010) have different takes on the nature of the problem. The treatment of the Prioniturus racket-tails is based on Schweitzer et al. (2012). They found that the race mindorensis is not part of P. discurus, but is more closely related to P. platenae. Both plumage and genetic distance suggest it is a separate species. I could find an existing English name, so I'm referring to it as Mindoro Racket-tail.
Psittacellini: Tiger-Parrots: Joseph et al. (2011) show that the Psittacellini tiger-parrots are sister to the remaining 4 tribes. Exactly how those 4 tribes fit together remains somewhat uncertain. I'm following the Schweizer et al. (2010) topology, which is consistent with the MP analysis of Wright et al. (2008), and figure 2 of Joseph et al. (2011). Joseph et al. argue that this result, from a restricted data set, is more likely to be correct. Their figure 1 and the Bayesian analysis in Wright et al. give a different arrangement.
Pezoporini: Ground and Night Parrots: This clade of distinctive parrots has been long separated from its closest relatives, which are likely the Platycercini. Their common ancestor may have lived in the Oligocene (Schweizer et al, 2011). Because of this, I do not follow Collar (1997) who includes them in the Platycercini.
Platycercini: Rosellas and allies: Platycercini has two parts: Psephotus through Platycercus and Lathamus through Cyanoramphus. All the genera of the first group are sometimes lumped in an expanded Platycercus. It appears that the traditional Psephotus is not monophyletic (see Joseph et al., 2011; Schweizer et al., 2010, 2013), with Red-rumped Parrot, Psephotus haematonotus, separated from the rest. As it is the type species for Psephotus, we go to the next available name for the other birds formerly placed in Psephotus. That name appears to be Psephotellus (Mathews 1913, type pulcherrimus). I'm following a blend of Schweizer et al. (2013) and Joseph et al. (2011) for this branch of the Platycercini. Although they are clearly part of the clade, the placement of Northiella and Psephotus remains uncertain. They show up in slightly different locations in different analyses.
Agapornithini: Lovebirds and Hanging-Parrots: The arrangement of Agapornis is based on Kundu et al. (2012).
Loriini: Fig-Parrots, Lories, and Lorikeets: At last we come to the Loriini. The Fig-Parrots, Cyclopsitta + Psittaculirostris, form the basal group in the Loriini. Then comes the budgie, Melopsittacus, followed by the lory/lorikeet clade. The division between the budgie and the lories/lorikeets is fairly deep.
Psittaculini Vigors, 1825
- Yellow-capped Pygmy-Parrot, Micropsitta keiensis
- Geelvink Pygmy-Parrot, Micropsitta geelvinkiana
- Buff-faced Pygmy-Parrot, Micropsitta pusio
- Meek's Pygmy-Parrot, Micropsitta meeki
- Finsch's Pygmy-Parrot, Micropsitta finschii
- Red-breasted Pygmy-Parrot, Micropsitta bruijnii
- Moluccan King-Parrot, Alisterus amboinensis
- Papuan King-Parrot, Alisterus chloropterus
- Australian King-Parrot, Alisterus scapularis
- Olive-shouldered Parrot / Jonquil Parrot, Aprosmictus jonquillaceus
- Red-winged Parrot, Aprosmictus erythropterus
- Superb Parrot, Polytelis swainsonii
- Regent Parrot, Polytelis anthopeplus
- Alexandra's Parrot / Princess Parrot, Polytelis alexandrae
- Golden-mantled Racket-tail, Prioniturus platurus
- Buru Racket-tail, Prioniturus mada
- Mindanao Racket-tail, Prioniturus waterstradti
- Montane Racket-tail, Prioniturus montanus
- Blue-headed Racket-tail, Prioniturus platenae
- Mindoro Racket-tail, Prioniturus mindorensis
- Blue-winged Racket-tail, Prioniturus verticalis
- Yellow-breasted Racket-tail, Prioniturus flavicans
- Green Racket-tail, Prioniturus luconensis
- Blue-crowned Racket-tail, Prioniturus discurus
- Eclectus Parrot, Eclectus roratus
- Red-cheeked Parrot, Geoffroyus geoffroyi
- Blue-collared Parrot, Geoffroyus simplex
- Song Parrot, Geoffroyus heteroclitus
- Blue-rumped Parrot, Psittinus cyanurus
- Great-billed Parrot, Tanygnathus megalorynchos
- Blue-naped Parrot, Tanygnathus lucionensis
- Blue-backed Parrot, Tanygnathus sumatranus
- Black-lored Parrot, Tanygnathus gramineus
- Broad-billed Parrot, Lophopsittacus mauritianus
- Rodrigues Parrot, Necropsittacus rodricanus
- Slaty-headed Parakeet, Psittacula himalayana
- Gray-headed Parakeet, Psittacula finschii
- Plum-headed Parakeet, Psittacula cyanocephala
- Blossom-headed Parakeet, Psittacula roseata
- Alexandrine Parakeet, Psittacula eupatria
- Seychelles Parakeet, Psittacula wardi
- Rose-ringed Parakeet, Psittacula krameri
- Echo Parakeet, Psittacula eques
- Mascarene Grey Parakeet, Psittacula bensoni
- Newton's Parakeet, Psittacula exsul
- Lord Derby's Parakeet, Psittacula derbiana
- Red-breasted Parakeet, Psittacula alexandri
- Nicobar Parakeet, Psittacula caniceps
- Long-tailed Parakeet, Psittacula longicauda
- Malabar Parakeet / Blue-winged Parakeet, Psittacula columboides
- Layard's Parakeet, Psittacula calthrapae
Psittacellini: Tiger-Parrots Wolters, 1975
Joseph et al. (2011) found that the Tiger-Parrots were not closely related to any of the remaining parrot tribes, but formed a branch sister to all of them.
- Brehm's Tiger-Parrot, Psittacella brehmii
- Painted Tiger-Parrot, Psittacella picta
- Modest Tiger-Parrot, Psittacella modesta
- Madarasz's Tiger-Parrot, Psittacella madaraszi
Pezoporini: Ground and Night Parrots Bonaparte, 1838
The Ground Parrot, Pezoporus wallicus, has been split into Western Ground Parrot, Pezoporus flaviventris, and Eastern Ground Parrot, Pezoporus wallicus. See Murphy et al. (2011).
- Night Parrot, Pezoporus occidentalis
- Western Ground Parrot, Pezoporus flaviventris
- Eastern Ground Parrot, Pezoporus wallicus
- Bourke's Parrot, Neopsephotus bourkii
- Blue-winged Parrot, Neophema chrysostoma
- Elegant Parrot, Neophema elegans
- Rock Parrot, Neophema petrophila
- Orange-bellied Parrot, Neophema chrysogaster
- Turquoise Parrot, Neophema pulchella
- Scarlet-chested Parrot, Neophema splendida
Platycercini Selby, 1836
The genus name Prosopeia (Bonaparte, 1854) has been replaced by Pyrrhulopsis (Reichenbach, 1850), which has priority. See Gregory and Dickinson (2012).
The Bluebonnet, Northiella haematogaster, has been split into Naretha Bluebonnet, Northiella narethae, and Eastern Bluebonnet, Northiella haematogaster. See Dolman and Joseph (2015).
The rosellas (Platycercus) have been rearranged based on Shipman et al. (2015). Even their multi-gene analysis was unable to pin down the position of the Western Rosella, Platycercus icterotis. It might the basal Platycercus, as shown here, or it might group with the white-cheeked species (eximius, venustus, adscitus).
- Red-rumped Parrot, Psephotus haematonotus
- Naretha Bluebonnet, Northiella narethae
- Eastern Bluebonnet, Northiella haematogaster
- Red-capped Parrot, Purpureicephalus spurius
- Mulga Parrot, Psephotellus varius
- Hooded Parrot, Psephotellus dissimilis
- Golden-shouldered Parrot, Psephotellus chrysopterygius
- Paradise Parrot, Psephotellus pulcherrimus
- Australian Ringneck, Barnardius zonarius
- Western Rosella, Platycercus icterotis
- Crimson Rosella, Platycercus elegans
- Green Rosella, Platycercus caledonicus
- Eastern Rosella, Platycercus eximius
- Northern Rosella, Platycercus venustus
- Pale-headed Rosella, Platycercus adscitus
- Swift Parrot, Lathamus discolor
- Crimson Shining-Parrot, Pyrrhulopsis splendens
- Masked Shining-Parrot, Pyrrhulopsis personatus
- Red Shining-Parrot / Maroon Shining-Parrot, Pyrrhulopsis tabuensis
- Horned Parakeet, Eunymphicus cornutus
- Ouvea Parakeet, Eunymphicus uvaeensis
- Black-fronted Parakeet, Cyanoramphus zealandicus
- Raiatea Parakeet, Cyanoramphus ulietanus
- New Caledonian Parakeet, Cyanoramphus saisseti
- Chatham Parakeet, Cyanoramphus forbesi
- Tasman Parakeet / Norfolk Parakeet, Cyanoramphus cookii
- Lord Howe Parakeet, Cyanoramphus subflavescens
- Antipodes Parakeet, Cyanoramphus unicolor
- Yellow-fronted Parakeet / Yellow-crowned Parakeet, Cyanoramphus auriceps
- Malherbe's Parakeet, Cyanoramphus malherbi
- Red-crowned Parakeet, Cyanoramphus novaezelandiae
- Reischek's Parakeet, Cyanoramphus hochstetteri
- Macquarie Parakeet, Cyanoramphus erythrotis
Agapornithini Salvin, 1882
The arrangement of Agapornis is based on Kundu et al. (2012). The available genetic information on the hanging-parrots is insufficient to order them, so I use a traditional ordering.
- Guaiabero, Bolbopsittacus lunulatus
- Red-headed Lovebird, Agapornis pullarius
- Black-winged Lovebird, Agapornis taranta
- Black-collared Lovebird, Agapornis swindernianus
- Gray-headed Lovebird, Agapornis canus
- Rosy-faced Lovebird, Agapornis roseicollis
- Fischer's Lovebird, Agapornis fischeri
- Yellow-collared Lovebird, Agapornis personatus
- Lilian's Lovebird / Nyasa Lovebird, Agapornis lilianae
- Black-cheeked Lovebird, Agapornis nigrigenis
- Vernal Hanging-Parrot, Loriculus vernalis
- Sri Lanka Hanging-Parrot, Loriculus beryllinus
- Colasisi / Philippine Hanging-Parrot, Loriculus philippensis
- Camiguin Hanging-Parrot, Loriculus camiguinensis
- Blue-crowned Hanging-Parrot, Loriculus galgulus
- Sulawesi Hanging-Parrot / Great Hanging-Parrot, Loriculus stigmatus
- Sula Hanging-Parrot, Loriculus sclateri
- Moluccan Hanging-Parrot, Loriculus amabilis
- Sangihe Hanging-Parrot, Loriculus catamene
- Orange-fronted Hanging-Parrot, Loriculus aurantiifrons
- Bismarck Hanging-Parrot, Loriculus tener
- Red-billed Hanging-Parrot / Pygmy Hanging-Parrot, Loriculus exilis
- Yellow-throated Hanging-Parrot, Loriculus pusillus
- Wallace's Hanging-Parrot, Loriculus flosculus
Loriini: Lories Selby, 1836
The name Coriphilus (Wagler, 1832) replaces Vini (Lesson, 1833, not 1831). Although the change in date for Vini seems correct and can be found in H&M-4 (Dickinson and Remsen, 2013), it doesn't seem to have been adopted. Because Coriphilus has been used since 1900 (at least up to the 1930s), it is impossible to use article 23.9 of the code to retain Vini. Hence I use Coriphilus.
Based on Schweizer et al. (2015), the Loriini have been reordered. Two of Glossopsitta have been separated in Parvipsitta (Mathews 1916, type pusilla). Goldie's Lorikeet moves from Psitteuteles to Glossoptilus (Rothschild and Hartert, 1896) as Glossoptilus goldiei. The Cardinal Lory joins Pseudeos as Pseudeos cardinalis (was Chalcopsitta) and the Iris Lory is now “Psitteuteles” iris. Note that the position of “Psitteuteles” is somewhat ambigious, so I left it in a trichotomy with Eos and Trichoglossus.
Schweizer et al. (2015) also found that Charmosyna is paraphyletic with respect to Coriphilus and Phigys, but did not sample enought taxa to be sure of how to handle Charmosyna. The tree doesn't fit comfortably with plumages, and its far from clear where several of the taxa will go. There might also be an issue with the Mindanao Lorikeet, Trichoglossus johnstoniae.
Now that Bird Life International has joined IOC in splitting the Rainbow Lorikeet, Trichoglossus haematodus, into three species, I'm adopting that split. BLI based their split is based on the Tobias et al. (2010) criteria for allopatric species, which is a reasonable rule of thumb in the absence of conflicting evidence. However, as BLI has not yet released a detailed analysis, I will not generally adopt these splits. In this case, I make an exception because IOC had already adopted them.
| Division of Rainbow Lorikeet | ||
|---|---|---|
| Coconut Lorikeet, Trichoglossus haematodus |
haematodus, intermedius, nigrogularis, caeruleiceps, micropteryx, massena, nesophilus, flavicans, and deplanchii | |
| Biak Lorikeet, Trichoglossus rosenbergii |
monotypic | |
| Rainbow Lorikeet, Trichoglossus moluccanus |
septentrionalis, moluccanus, and eyrei | |
- Orange-breasted Fig-Parrot, Cyclopsitta gulielmitertii
- Double-eyed Fig-Parrot, Cyclopsitta diophthalma
- Large Fig-Parrot, Psittaculirostris desmarestii
- Edwards's Fig-Parrot, Psittaculirostris edwardsii
- Salvadori's Fig-Parrot, Psittaculirostris salvadorii
- Budgerigar, Melopsittacus undulatus
- Plum-faced Lorikeet, Oreopsittacus arfaki
- Collared Lory, Phigys solitarius
- Blue-crowned Lorikeet, Coriphilus australis
- Kuhl's Lorikeet, Coriphilus kuhlii
- Stephen's Lorikeet / Henderson Lorikeet, Coriphilus stepheni
- Blue Lorikeet, Coriphilus peruvianus
- Ultramarine Lorikeet, Coriphilus ultramarinus
- Palm Lorikeet, Charmosyna palmarum
- Red-chinned Lorikeet, Charmosyna rubrigularis
- Meek's Lorikeet, Charmosyna meeki
- Blue-fronted Lorikeet, Charmosyna toxopei
- Striated Lorikeet, Charmosyna multistriata
- Pygmy Lorikeet, Charmosyna wilhelminae
- Red-fronted Lorikeet, Charmosyna rubronotata
- Red-flanked Lorikeet, Charmosyna placentis
- New Caledonian Lorikeet, Charmosyna diadema
- Red-throated Lorikeet, Charmosyna amabilis
- Duchess Lorikeet, Charmosyna margarethae
- Fairy Lorikeet, Charmosyna pulchella
- Josephine's Lorikeet, Charmosyna josefinae
- Papuan Lorikeet, Charmosyna papou
- Yellow-billed Lorikeet, Neopsittacus musschenbroekii
- Orange-billed Lorikeet, Neopsittacus pullicauda
- Little Lorikeet, Parvipsitta pusilla
- Purple-crowned Lorikeet, Parvipsitta porphyrocephala
- Chattering Lory, Lorius garrulus
- Purple-naped Lory, Lorius domicella
- Black-capped Lory, Lorius lory
- Purple-bellied Lory, Lorius hypoinochrous
- White-naped Lory, Lorius albidinucha
- Yellow-bibbed Lory, Lorius chlorocercus
- Varied Lorikeet, Psitteuteles versicolor
- Goldie's Lorikeet, Glossoptilus goldiei
- Iris Lorikeet, "Psitteuteles" iris
- Dusky Lory, Pseudeos fuscata
- Cardinal Lory, Pseudeos cardinalis
- Brown Lory, Chalcopsitta duivenbodei
- Black Lory, Chalcopsitta atra
- Yellowish-streaked Lory, Chalcopsitta scintillata
- Musk Lorikeet, Glossopsitta concinna
- Blue-streaked Lory, Eos reticulata
- Black-winged Lory, Eos cyanogenia
- Red-and-blue Lory, Eos histrio
- Violet-necked Lory, Eos squamata
- Red Lory, Eos bornea
- Blue-eared Lory, Eos semilarvata
- Ornate Lorikeet, Trichoglossus ornatus
- Scarlet-breasted Lorikeet / Sunset Lorikeet, Trichoglossus forsteni
- Flores Lorikeet / Leaf Lorikeet, Trichoglossus weberi
- Marigold Lorikeet, Trichoglossus capistratus
- Coconut Lorikeet, Trichoglossus haematodus
- Biak Lorikeet, Trichoglossus rosenbergii
- Rainbow Lorikeet, Trichoglossus moluccanus
- Red-collared Lorikeet, Trichoglossus rubritorquis
- Olive-headed Lorikeet, Trichoglossus euteles
- Yellow-and-green Lorikeet / Citrine Lorikeet, Trichoglossus flavoviridis
- Mindanao Lorikeet, Trichoglossus johnstoniae
- Pohnpei Lorikeet, Trichoglossus rubiginosus
- Scaly-breasted Lorikeet, Trichoglossus chlorolepidotus