GALLIFORMES Temminck, 1820
The Galliformes are sister to the Anseriformes. Together, they form the Galloanserae, the sister group to Neoaves.
The split between the Anseriformes and the Galliformes is estimated to have occurred 62.52 mya (64.9–56.8 mya) by Kuhl et al. (2021) and 68.1±2.2mya by Stiller et al. (2024).
 |
| Click for genus-level tree for Galliformes |
|---|
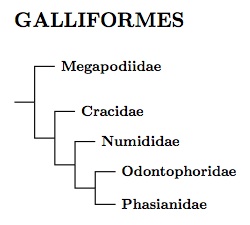
The arrangement of Galliform families follows the genetic analyses of Crowe et al. (2006a), Cox et al. (2007), Hackett et al. (2008), and Meiklejohn et al. (2014). It is also consistent with Kaiser et al. (2007) and Kriegs et al. (2007), who analyze retropositions. Of the two, Kriegs et al. is more complete, and exhibits the same basic structure as the tree here. Since it is a different method of reading the genetic data, it represents significant additional support for the current arrangement of Galliform families. Nonetheless, there is still some question about whether the Numidade branch off before or after the Odontophoridae (e.g., Shen et al., 2010).
The Galliform tree takes the form of a cascade, with one group breaking off at a time until you get to the Phasianidae. It starts with the megapodes, then the Cracidae (guans, chachalacas, and curassows), guineafowl, and finally New World Quail, all before we get to the Phasianidae. A number of genera have been rearranged or split compared with previous treatments. The genera decorated with question marks are uncertainly placed and could easily end up in a different subfamily.
Two of the Galliform lineages are quite ancient. Based on Fig. ED-2, Stiller et al.'s point estimate of the Megapodiidae split is 66.8 mya, shortly before the end of the Cretaceous period. They show the Cracidae split dating to around 59.5 mya. Both are old enough to deserve consideration as separate orders. However, although they are definitely different from the other Galliformes (and each other), these families don't seem really out of place in the Galliformes. I may come back to this later, but for now (2024), I don't feel compelled to set up two more orders for them (Megapodiiformes and Craciformes).
Megapodiidae: Megapodes Lesson, 1831
7 genera, 22 species HBW-2
The arrangement of the megapodes is based on the complete multi-gene analysis by Harris et al. (2014b), which is similiar to the sparser 2-gene analysis of Birks and Edwards (2002).
- Collared Brush-turkey, Talegalla jobiensis
Click for Megapodiidae
species tree - Red-billed Brush-turkey, Talegalla cuvieri
- Black-billed Brush-turkey, Talegalla fuscirostris
- Malleefowl, Leipoa ocellata
- Australian Brush-turkey, Alectura lathami
- Waigeo Brush-turkey, Aepypodius bruijnii
- Wattled Brush-turkey, Aepypodius arfakianus
- Maleo, Macrocephalon maleo
- Moluccan Megapode, Eulipoa wallacei
- Philippine Megapode, Megapodius cumingii
- Tanimbar Megapode, Megapodius tenimberensis
- Nicobar Megapode, Megapodius nicobariensis
- Micronesian Megapode, Megapodius laperouse
- Vanuatu Megapode, Megapodius layardi
- Tongan Megapode, Megapodius pritchardii
- Sula Megapode, Megapodius bernsteinii
- Biak Scrubfowl, Megapodius geelvinkianus
- Orange-footed Scrubfowl, Megapodius reinwardt
- Melanesian Megapode, Megapodius eremita
- New Guinea Scrubfowl, Megapodius decollatus
- Dusky Megapode, Megapodius freycinet
- Forsten's Megapode, Megapodius forsteni
Cracidae: Chachalacas, Curassows, Guans Rafinesque, 1815
9 genera, 54 species HBW-2
There has been a debate concerning whether the chachalacas and Horned Guan are more closely related to the guans or currasows. Pereira et al. (2002, 2009) found that the Horned Guan is closer to the currasows, and that the chachalacas are sister to the Horned Guan-Currasow clade. This topology has generally been supported by other studies, e.g., Kimball et al. (2011), although Frank-Hoeflich et al. (2006) found the opposite branching order using combined molecular, osteological, morphological, and behaviorial data (but see the response by Pereira et al., 2009).
Pereira et al. (2002) estimated divergence times for various genus-level nodes. They found several divisions dating back to the Oligocene. I've used subfamilies and tribes to highlight these divisions. Except for the Horned Guan, they correspond to the traditional division into chachalacas, curassows, and guans.
The genus Pipile has been submerged into Aburria (Grau et al., 2005). The arrangement of Aburria is based on Grau et al. (2005) and Robinson (2011).
No comprehensive study of the guans is available. The order follows SACC. The chachalaca ordering is based on the analyses of Frank-Hoeflich et al. (2006). Interestingly, they found the Ortalis canicollis race pantanalensis on a separate branch from O. canicollis, whether this is an artifact or real is unclear. The order within the currasows is based on Pereira and Baker (2004), which also requires merging Mitu into Pauxi.
Based on Gastañaga et al. (2011), Sira Curassow, Pauxi koepckeae, from the Sira Mountains in Peru, has been split from Horned Curassow, Pauxi unicornis.
The SACC has split East Brazilian Chachalaca, Ortalis araucuan, and Scaled Chachalaca, Ortalis squamata, from Speckled Chachalaca, Ortalis guttata.
Penelopinae: Guans Bonaparte, 1831
- Black Guan, Chamaepetes unicolor
- Sickle-winged Guan, Chamaepetes goudotii
- Highland Guan, Penelopina nigra
- Black-fronted Piping-Guan, Aburria jacutinga
- Trinidad Piping-Guan, Aburria pipile
- Wattled Guan, Aburria aburri
- Blue-throated Piping-Guan, Aburria cumanensis
- Red-throated Piping-Guan, Aburria cujubi
- Band-tailed Guan, Penelope argyrotis
- Bearded Guan, Penelope barbata
- Baudo Guan, Penelope ortoni
- Andean Guan, Penelope montagnii
- Marail Guan, Penelope marail
- Rusty-margined Guan, Penelope superciliaris
- Red-faced Guan, Penelope dabbenei
- Spix's Guan, Penelope jacquacu
- Crested Guan, Penelope purpurascens
- Cauca Guan, Penelope perspicax
- White-winged Guan, Penelope albipennis
- Dusky-legged Guan, Penelope obscura
- White-crested Guan, Penelope pileata
- Chestnut-bellied Guan, Penelope ochrogaster
- White-browed Guan, Penelope jacucaca
Cracinae: Chachalacas and Curassows Rafinesque, 1815
Various issues have arisen concerning the Chachalaca tribe. I now use Donegan's Ortalidaini (Donegan, 2012) as all more natural forms are preoccupied by names based on the fly genus Ortalis Fallén 1810, a junior homonym of the chachalaca genus Ortalis Merrem 1786. The genus Ortalis Fannén 1810 has been used to establish the family-group names Ortalides Fallén, 1810, Ortalidae Swainson 1840, Ortalididae sensu Harris 1841, and Ortalisini Acloque, 1897, all of which are based on Ortalis Fallén 1810. See Donegan (2015). The last case rules out use of David's (2014) correction of Donegan's spelling Ortalidaini. There is also a discrepancy between the French and English versions of the ICXN that calls into question whether Ortalidaini is properly formed. Still, it is the best current option pending the outcome of ICZN case 3669 (Donegan, 2015).
Ortalidaini: Chachalacas Donegan, 2012
- Variable Chachalaca / Little Chachalaca, Ortalis motmot
- White-bellied Chachalaca, Ortalis leucogastra
- Plain Chachalaca, Ortalis vetula
- Rufous-bellied Chachalaca, Ortalis wagleri
- West Mexican Chachalaca, Ortalis poliocephala
- Rufous-vented Chachalaca, Ortalis ruficauda
- Gray-headed Chachalaca, Ortalis cinereiceps
- Chestnut-winged Chachalaca, Ortalis garrula
- Rufous-headed Chachalaca, Ortalis erythroptera
- Chaco Chachalaca, Ortalis canicollis
- Colombian Chachalaca, Ortalis columbiana
- Speckled Chachalaca, Ortalis guttata
- East Brazilian Chachalaca, Ortalis araucuan
- Scaled Chachalaca, Ortalis squamata
- Buff-browed Chachalaca, Ortalis superciliaris
Oreophaseini: Horned Guan Bonaparte, 1853
- Horned Guan (Oreophasis derbianus)
Cracini: Curassows Rafinesque, 1815
- Nocturnal Curassow, Nothocrax urumutum
- Helmeted Curassow, Pauxi pauxi
- Crestless Curassow, Pauxi tomentosa
- Alagoas Curassow, Pauxi mitu
- Salvin's Curassow, Pauxi salvini
- Sira Curassow, Pauxi koepckeae
- Horned Curassow, Pauxi unicornis
- Razor-billed Curassow, Pauxi tuberosa
- Great Curassow, Crax rubra
- Blue-billed Curassow, Crax alberti
- Yellow-knobbed Curassow, Crax daubentoni
- Wattled Curassow, Crax globulosa
- Red-billed Curassow, Crax blumenbachii
- Black Curassow, Crax alector
- Bare-faced Curassow, Crax fasciolata
Numididae: Guineafowl de Selys Longchamps, 1842
4 genera, 6 species HBW-2
The order of genera follows Crowe et al. (2006a, b). This coincides with the cytochrome-b tree in Crowe et al. (2006a). Kimball et al. (2011) give a slighty different arrangement that swaps Numida and Guttera.
- Plumed Guineafowl, Guttera plumifera
- Crested Guineafowl, Guttera pucherani
- Helmeted Guineafowl, Numida meleagris
- Vulturine Guineafowl, Acryllium vulturinum
- White-breasted Guineafowl, Agelastes meleagrides
- Black Guineafowl, Agelastes niger
Odontophoridae: New World Quail Gould, 1844
11 genera, 34 species HBW-2
Surprisingly, two Old World species thought to be unrelated members of Phasianidae have turned out to be sister taxa and basal members of the Odontophoridae. They are the Stone Partridge (Ptilopachus petrosus) and Nahan's Francolin, which I've moved to Acentrortyx (Chapin, 1926) as Nahan's Partridge, Acentrortyx nahani. See Crowe et al. (2006a) and Cohen et al. (2012). Although Cohen et al. recommend including Nahan'a Partridge in Ptilopachus, it has long been considered to belong to a separate genus and lives in an entirely different habitat. Moreover, they estimate a divergence time of 9.6 mya between the two taxa. The chronogram in Hosner et al. (2015a) makes clear that all other such divergences in Odontophoridae are treated in separate genera. Taking all this into account, I put the two African partridges in separate genera.
The division between the two African partridges and the New World Quail is quite ancient. Cohen et al. (2012) estimated it between 33.9 and 55.8 mya (most likely in the Eocene), while Hosner et al. (2015a) put it between 27 and 38 mya (mid Oligocene to late-Eocene). Accordingly, I treat them as separate subfamilies. Bowie et al. (2013) created the name Ptilopachinae for this.
The New World Quail are arranged based on Hosner et al. (2015a). Different methods of estimation gave slightly different results for Odontophorus. The biggest change involved the Spotted Wood-Quail, Odontophorus guttatus. Their concatenated tree put it sister to Stripe-faced Wood-Quail, Odontophorus balliviani. Their coalescent tree (Fig. 3b) is followed here.
Ptilopachinae: Stone and Nahan's Partridges Bowie, Coehn, & Crowe, 2013
- Stone Partridge, Ptilopachus petrosus
- Nahan's Partridge, Acentrortyx nahani
Odontophorinae: New World Quail Gould, 1844
- Tawny-faced Quail, Rhynchortyx cinctus
Click for Odontophoridae
species tree - Mountain Quail, Oreortyx pictus
- Bearded Wood-Partridge, Dendrortyx barbatus
- Long-tailed Wood-Partridge, Dendrortyx macroura
- Buffy-crowned Wood-Partridge, Dendrortyx leucophrys
- Banded Quail, Philortyx fasciatus
- California Quail, Callipepla californica
- Gambel's Quail, Callipepla gambelii
- Scaled Quail, Callipepla squamata
- Elegant Quail, Callipepla douglasii
- Crested Bobwhite, Colinus cristatus
- Spot-bellied Bobwhite, Colinus leucopogon
- Northern Bobwhite, Colinus virginianus
- Black-throated Bobwhite / Yucatan Bobwhite, Colinus nigrogularis
- Montezuma Quail, Cyrtonyx montezumae
- Ocellated Quail, Cyrtonyx ocellatus
- Singing Quail, Dactylortyx thoracicus
- Spotted Wood-Quail, Odontophorus guttatus
- Marbled Wood-Quail, Odontophorus gujanensis
- Black-eared Wood-Quail, Odontophorus melanotis
- Rufous-fronted Wood-Quail, Odontophorus erythrops
- Starred Wood-Quail, Odontophorus stellatus
- Spot-winged Wood-Quail, Odontophorus capueira
- Stripe-faced Wood-Quail, Odontophorus balliviani
- Chestnut Wood-Quail, Odontophorus hyperythrus
- Dark-backed Wood-Quail, Odontophorus melanonotus
- Rufous-breasted Wood-Quail, Odontophorus speciosus
- Black-fronted Wood-Quail, Odontophorus atrifrons
- Tacarcuna Wood-Quail, Odontophorus dialeucos
- Gorgeted Wood-Quail, Odontophorus strophium
- Venezuelan Wood-Quail, Odontophorus columbianus
- Black-breasted Wood-Quail, Odontophorus leucolaemus
Phasianidae: Turkeys, Grouse, Pheasants, Partridges Horsfield, 1821
54 genera, 184 species HBW-2
The divisions within the Phasianidae remain a problem. The papers by Bao et al. (2010), Bonilla et al. (2010), Crowe et al. (2006a, b), Kan et al. (2010), Kimball and Braun (2008), Kimball et al. (2011), Kriegs et al. (2007), Liu et al. (2012) Meng et al. (2008), and Shen et al. (2010) give different answers about how the various components of Phasianidae are related. This is especially true if you consider the individual gene trees contained in Shen et al.'s supplementary material. And what is one to make of Liu et al. (2012)?
There does seem to be a broad, but not complete consensus that the Asiatic partridges (Rollulinae) are basal (see Liu et al. (2012) for a contrary view), and that some group containing Gallus is sister to a group containing the Phasianinae, but the placement of various pheasants (Pavonini, and Polyplectronini) and even the placement of the Tetraogallini is problematic (ditto its internal organization). Sometimes these tribes group near the Gallini in various configurations, sometimes they are not in a clade with Gallini and Phasianinae. Right now, the consensus seems to be running in favor of grouping these four tribes together (Pavonini, Polyplectronini, Gallini, and Tetraogallini) as Pavoninae but leave the branching order unresolved for the present. They are listed in order of size.
How to piece together the Phasianinae subfamily had been a little tricky, but a consistent picture has now emerged. The current order is based on Kimball et al. (2011), which includes most of the species in the subfamily. Other than including more taxa, it is quite similar to the results of Kimball and Braun (2008). Phasianinae is their “erectile clade”. I have divided it into tribes to better show the relationships of the various groups. As you can see by their membership, these tribes generally correspond to what we think of as natural groupings. Bao et al. (2010) is generally consistent with the present arrangement of the Phasianinae. Crowe et al. (2006a) suggested that the turkeys and Perdix partridges might be sister genera, but that has gotten little support from recent analyses.
Not surprisingly, part of the reconstruction of the Phasianidae involved moving some of the species to new genera. Crowe et al. (2006a) found two species that were quite wrongly placed: the Stone Partridge (Ptilopachus petrosus) and Nahan's Francolin (now Nahan's Partridge, Ptilopachus nahani), both of which ended up in the New World Quail (Odontophoridae). When Crowe et al. (1992, with a different set of co-authors) had reorganized the francolins, they noted that Nahan's Francolin didn't appear to be a francolin. That reorganization is mostly supported by the new paper, but one other francolin proved problematic. The Crested Francolin, which they had already reassigned to the genus Peliperdix, is now Dendroperdix sephaena, although it remains in the junglefowl/francolin subfamily, Gallini.
Eo et al. (2009) draws attention to several genera that are currently regarded as part of Phasianidae (Haematortyx, Melanoperdix, Rhizothera, Galloperdix). While it suggests closer scrutiny of these genera is in order, I don't buy the arrangement of them in the paper. I think it's an artifact of the supertree method. In fact, Sun et al. (2014) found that Haematortyx and Galloperdix belong near Polyplectron, a very different location than in Eo et al. Crowe et al. (2006a), does find that the Ptilopachus-Odontophoridae clade is sister to the Phasianidae, which is why I've moved Ptilopachus into Odontophoridae. They did not find Phasianidae nested within the Ptilopachus-Odontophoridae clade. I'm leaving the other genera highlighted by Eo et al. in Phasianidae for now, but we should keep in mind that some or all may too belong in Odontophoridae.
Rollulinae: Asiatic Partridges Bonaparte, 1850 (1848)
Bonaparte's name Rollulinae has priority over Arborophilinae, Crowe et al. (2006a). Note that the Rubeho Forest Partridge, Xenoperdix obscuratus has been split from Udzungwa Forest Partridge, Xenoperdix udzungwensis (Bowie and Fjeldså, 2005). See Wang et al. (2013) for the treatment of Caloperdix as sister to Rollulus.
- Rubeho Forest Partridge, Xenoperdix obscuratus
- Udzungwa Forest Partridge, Xenoperdix udzungwensis
- Black Partridge, Melanoperdix niger
- Long-billed Partridge, Rhizothera longirostris
- Hose's Partridge, Rhizothera dulitensis
- Ferruginous Partridge, Caloperdix oculeus
- Crested Partridge, Rollulus rouloul
- Hill Partridge, Arborophila torqueola
- Rufous-throated Partridge, Arborophila rufogularis
- White-cheeked Partridge, Arborophila atrogularis
- Taiwan Partridge, Arborophila crudigularis
- Chestnut-breasted Partridge, Arborophila mandellii
- Bar-backed Partridge, Arborophila brunneopectus
- Sichuan Partridge, Arborophila rufipectus
- White-necklaced Partridge, Arborophila gingica
- Orange-necked Partridge, Arborophila davidi
- Chestnut-headed Partridge, Arborophila cambodiana
- Siamese Partridge, Arborophila diversa
- Malaysian Partridge, Arborophila campbelli
- Roll's Partridge, Arborophila rolli
- Sumatran Partridge, Arborophila sumatrana
- Gray-breasted Partridge, Arborophila orientalis
- Chestnut-bellied Partridge, Arborophila javanica
- Red-billed Partridge, Arborophila rubrirostris
- Red-breasted Partridge, Arborophila hyperythra
- Hainan Partridge, Arborophila ardens
Pavoninae: Peafowl and allies Rafinesque, 1815
Incertae Sedis: Pavoninae
The Chestnut-necklaced Partridge and Green-legged Partridge have been moved out of Arborophila and Rollulinae to become genus Tropicoperdix in Pavoninae. It is uncertain exactly where they belong in Pavoninae. One possibility is that they are basal in Pavonini. For now, they are left incertae sedis in Pavoninae (Chen et al., 2015).
- Green-legged Partridge, Tropicoperdix chloropus
Click for Pavoninae
species tree - Chestnut-necklaced Partridge, Tropicoperdix charltonii
Pavonini: Peafowl Rafinesque, 1815
- Great Argus, Argusianus argus
- Crested Argus, Rheinardia ocellata
- Congo Peafowl, Afropavo congensis
- Indian Peafowl, Pavo cristatus
- Green Peafowl, Pavo muticus
Polyplectronini: Peacock-Pheasants Blyth, 1852
The arrangement of Polyplectron is based on Davison et al. (2012), Kimball et al. (2011, 2014) and Sun et al. (2014). Sun et al. (2014 also found that Galloperdix belongs here.
- Crimson-headed Partridge, Haematortyx sanguiniceps
- Red Spurfowl, Galloperdix spadicea
- Painted Spurfowl, Galloperdix lunulata
- Sri Lanka Spurfowl, Galloperdix bicalcarata
- Palawan Peacock-Pheasant, Polyplectron napoleonis
- Malayan Peacock-Pheasant, Polyplectron malacense
- Bornean Peacock-Pheasant, Polyplectron schleiermacheri
- Germain's Peacock-Pheasant, Polyplectron germaini
- Hainan Peacock-Pheasant, Polyplectron katsumatae
- Mountain Peacock-Pheasant, Polyplectron inopinatum
- Gray Peacock-Pheasant, Polyplectron bicalcaratum
- Bronze-tailed Peacock-Pheasant, Polyplectron chalcurum
Gallini: Junglefowl, Francolins Brehm, 1831
Latham's Francolin, Peliperdix lathami, takes the basal position based on Kimball et al. (2011). The other 3 species sometimes placed in Peliperdix (coqui, albogularis, schlegelii) seem more closely related to the Scleroptila francolins, and are merged into that genus. Although the Crested Francolin, Francolinus sephaena, has been included in Peliperdix, indications are that it is in the francolin group (Crowe et al., 2006; Meng et al., 2008) perhaps closer to the Gray Francolin, Francolinus pondicerianus, than to the francolinus group (Kimball and Braun, 2011). The arrangement within the rest of Francolinus follows Forcina et al. (2012)
The Taiwan Bamboo-Partridge, Bambusicola sonorivox, has been split from the Chinese Bamboo-Partridge, Bambusicola thoracicus. See Hung et al. (2014).
The Orange River Francolin, Scleroptila gutturalis, has been split into Archer's Francolin (including subspecies archeri and lorti) and Orange River Francolin, Scleroptila levalliantoides. See Mandiwana-Neudani et al. (2014) and Sinclair and Ryan (2003). There are some other still-unpublished possible splits in Mandiwana-Neudani (2013).
- Mountain Bamboo-Partridge, Bambusicola fytchii
- Chinese Bamboo-Partridge, Bambusicola thoracicus
- Taiwan Bamboo-Partridge, Bambusicola sonorivox
- Red Junglefowl, Gallus gallus
- Green Junglefowl, Gallus varius
- Gray Junglefowl, Gallus sonneratii
- Sri Lanka Junglefowl, Gallus lafayettii
- Latham's Francolin, Peliperdix lathami
- Crested Francolin, Francolinus sephaena
- Gray Francolin, Francolinus pondicerianus
- Swamp Francolin, Francolinus gularis
- Chinese Francolin, Francolinus pintadeanus
- Black Francolin, Francolinus francolinus
- Painted Francolin, Francolinus pictus
- Coqui Francolin, Scleroptila coqui
- White-throated Francolin, Scleroptila albogularis
- Schlegel's Francolin, Scleroptila schlegelii
- Ring-necked Francolin, Scleroptila streptophora
- Red-winged Francolin, Scleroptila levaillantii
- Finsch's Francolin, Scleroptila finschi
- Moorland Francolin, Scleroptila psilolaema
- Archer's Francolin, Scleroptila gutturalis
- Shelley's Francolin, Scleroptila shelleyi
- Orange River Francolin, Scleroptila levalliantoides
- Gray-winged Francolin, Scleroptila afra
Tetraogallini: Old World Partridges and Quail, Spurfowl Bonaparte, 1854 (1846)
The Brown Quail is not closely related to the other Coturnix species (Seabrook-Davison et al., 2009; Kimball et al., 2011). Its closest relatives are Excalfactoria. However, it seems to be a fairly distant relative and I have placed it in Synoicus (Gould, 1843). Wang et al. (2013) found that Ammoperdix is a basal member of the Coturnix clade.
The arrangement within Pternistis is based on Mandiwana-Neudani et al. (2014).
Töpfer et al. (2014) recommend treating the Black-fronted Francolin, Pternistis castaneicollis atrifrons, as a separate species. However, the genetic differences are modest and I think more evidence is required.
- See-see Partridge, Ammoperdix griseogularis
- Sand Partridge, Ammoperdix heyi
- Brown Quail, Synoicus ypsilophorus
- King Quail, Excalfactoria chinensis
- Blue Quail, Excalfactoria adansonii
- Madagascan Partridge, Margaroperdix madagarensis
- Snow Mountain Quail, Anurophasis monorthonyx
- Common Quail, Coturnix coturnix
- Japanese Quail, Coturnix japonica
- Rain Quail, Coturnix coromandelica
- Harlequin Quail, Coturnix delegorguei
- Stubble Quail, Coturnix pectoralis
- New Zealand Quail, Coturnix novaezelandiae
- Caucasian Snowcock, Tetraogallus caucasicus
- Caspian Snowcock, Tetraogallus caspius
- Himalayan Snowcock, Tetraogallus himalayensis
- Tibetan Snowcock, Tetraogallus tibetanus
- Altai Snowcock, Tetraogallus altaicus
- Barbary Partridge, Alectoris barbara
- Arabian Partridge, Alectoris melanocephala
- Rock Partridge, Alectoris graeca
- Red-legged Partridge, Alectoris rufa
- Chukar / Chukar Partridge, Alectoris chukar
- Philby's Partridge, Alectoris philbyi
- Przevalski's Partridge, Alectoris magna
- Himalayan Quail, Ophrysia superciliosa
- Jungle Bush-Quail, Perdicula asiatica
- Rock Bush-Quail, Perdicula argoondah
- Painted Bush-Quail, Perdicula erythrorhyncha
- Manipur Bush-Quail, Perdicula manipurensis
- Hartlaub's Spurfowl, Pternistis hartlaubi
- Mount Cameroon Francolin, Pternistis camerunensis
- Handsome Francolin, Pternistis nobilis
- Chestnut-naped Francolin, Pternistis castaneicollis
- Erckel's Francolin, Pternistis erckelii
- Djibouti Francolin, Pternistis ochropectus
- Double-spurred Francolin, Pternistis bicalcaratus
- Heuglin's Francolin, Pternistis icterorhynchus
- Gray-striped Francolin, Pternistis griseostriatus
- Ahanta Francolin, Pternistis ahantensis
- Scaly Francolin, Pternistis squamatus
- Red-billed Spurfowl, Pternistis adspersus
- Cape Spurfowl, Pternistis capensis
- Hildebrandt's Francolin, Pternistis hildebrandti
- Natal Spurfowl, Pternistis natalensis
- Jackson's Francolin, Pternistis jacksoni
- Swierstra's Francolin, Pternistis swierstrai
- Clapperton's Francolin, Pternistis clappertoni
- Harwood's Francolin, Pternistis harwoodi
- Swainson's Spurfowl, Pternistis swainsonii
- Yellow-necked Spurfowl, Pternistis leucoscepus
- Red-necked Spurfowl, Pternistis afer
- Gray-breasted Spurfowl, Pternistis rufopictus
Phasianinae: Pheasants, Grouse, Turkeys Horsfield, 1821
The Vietnamese Pheasant, Lophura hatinhensis, is considered part of Edwards's Pheasant, Lophura edwardsi. It appears to be either a subspecies or color morph. In any event, it is not genetically distinct (Hennache et al., 2003). Also, the arrangement within Lophura is based on Randi et al., 2001.
Ithaginini: Blood Pheasant Wolters, 1976
- Blood Pheasant, Ithaginis cruentus
Click for Phasianinae
species tree
Lophophorini: Monals and Tragopans G.R. Gray, 1841
- Western Tragopan, Tragopan melanocephalus
- Satyr Tragopan, Tragopan satyra
- Blyth's Tragopan, Tragopan blythii
- Temminck's Tragopan, Tragopan temminckii
- Cabot's Tragopan, Tragopan caboti
- Snow Partridge, Lerwa lerwa
- Verreaux's Monal-Partridge, Tetraophasis obscurus
- Szechenyi's Monal-Partridge, Tetraophasis szechenyii
- Chinese Monal, Lophophorus lhuysii
- Himalayan Monal, Lophophorus impejanus
- Sclater's Monal, Lophophorus sclateri
Tetraonini: Grouse and Turkeys Leach, 1820
The Turkeys are now consiered embedded in Tetraonini. Interestingly enough, using a combined mitochondrial and nuclear dataset, Kimball and Braun (2014) found that the Koklass Pheasant, Pucrasia macrolopha, is basal in this group. Note that an analysis of the complete mitochondrial genome puts it basal in the Phasianini (Meiklejohn et al., 2014).
- Koklass Pheasant, Pucrasia macrolopha
- Wild Turkey, Meleagris gallopavo
- Ocellated Turkey, Meleagris ocellata
- Ruffed Grouse, Bonasa umbellus
- Hazel Grouse, Tetrastes bonasia
- Chinese Grouse, Tetrastes sewerzowi
- Greater Sage-Grouse / Sage Grouse, Centrocercus urophasianus
- Gunnison Sage-Grouse / Gunnison Grouse, Centrocercus minimus
- Dusky Grouse, Dendragapus obscurus
- Sooty Grouse, Dendragapus fuliginosus
- Sharp-tailed Grouse, Tympanuchus phasianellus
- Greater Prairie-Chicken, Tympanuchus cupido
- Lesser Prairie-Chicken, Tympanuchus pallidicinctus
- White-tailed Ptarmigan, Lagopus leucura
- Rock Ptarmigan, Lagopus muta
- Willow Ptarmigan / Red Grouse, Lagopus lagopus
- Siberian Grouse, Falcipennis falcipennis
- Spruce Grouse, Canachites canadensis
- Western Capercaillie, Tetrao urogallus
- Black-billed Capercaillie, Tetrao urogalloides
- Black Grouse, Lyrurus tetrix
- Caucasian Grouse, Lyrurus mlokosiewiczi
Phasianini: Pheasants, Perdix Partridges Horsfield, 1821
- Tibetan Partridge, Perdix hodgsoniae
- Gray Partridge, Perdix perdix
- Daurian Partridge, Perdix dauurica
- Reeves's Pheasant, Syrmaticus reevesii
- Copper Pheasant, Syrmaticus soemmerringii
- Mikado Pheasant, Syrmaticus mikado
- Elliot's Pheasant, Syrmaticus ellioti
- Mrs. Hume's Pheasant, Syrmaticus humiae
- Ring-necked Pheasant / Common Pheasant, Phasianus colchicus
- Green Pheasant, Phasianus versicolor
- Golden Pheasant, Chrysolophus pictus
- Lady Amherst's Pheasant, Chrysolophus amherstiae
- Cheer Pheasant, Catreus wallichii
- White Eared-Pheasant, Crossoptilon crossoptilon
- Tibetan Eared-Pheasant, Crossoptilon harmani
- Brown Eared-Pheasant, Crossoptilon mantchuricum
- Blue Eared-Pheasant, Crossoptilon auritum
- Siamese Fireback, Lophura diardi
- Crested Fireback, Lophura ignita
- Crestless Fireback, Lophura erythrophthalma
- Hoogerwerf's Pheasant, Lophura hoogerwerfi
- Salvadori's Pheasant, Lophura inornata
- Bulwer's Pheasant, Lophura bulweri
- Kalij Pheasant, Lophura leucomelanos
- Silver Pheasant, Lophura nycthemera
- Swinhoe's Pheasant, Lophura swinhoii
- Edwards's Pheasant, Lophura edwardsi