Passerida Nitzsch, 1820
Today, we have a reasonably good consensus on passerine phylogney. The TiF list now uses three large-scale studies: Oliveros et al. (2019), Kuhl et al. (2021), and Stiller et al. (2024). Oliveros et al. was the only one that sampled all of the families in Passeriformes, although it looked at fewer genes. Kuhl et al. and especially Stiller et al. sampled much more of the genome than Oliveros et al. Among the basal Passerida families, neither Kuhl et al. nor Stiller et al. included the rail-babbler. Kuhl et al. also left out the stitchbird and New Zealand wattlebirds.
The Passerida are the sister group of the Corvida. The two groups split in the mid-Oligocene, probably around 31 mya. Recent age estimates include 30.5 mya from Oliveros et al. (2019), 33.8 mya from Kuhl et al. (2021), and 29.3 mya from Stiller et al. (2024).
Passerida contains over 4000 species, more than a third of all bird species. As for the structure of the group, Passerida starts with several basal groups before we get to the heart of the Passerida. These are all ranked as superfamilies, and contain 1-4 families each. The early branches are Cnemophiloidea, Melanocharitoidea, Callaeoidea, and Eupetoidea.
Cnemophiloidea contains a single family, the satinbirds. They had previously been considered birds-of-paradise and are endemic to New Guinea. Melanocharitoidea split next. It also contains a single family, Melanocharitidae, berrypeckers and longbills. They are also endemic to New Guinea. The third superfamily, Callaeoidea, contains two families, Notiomystidae, the stitchbird, and Callaeidae, the New Zealand Wattlebirds. Both of these small families are endemic to New Zealand.
The last small superfamily is Eupetoidea. This group is also likely of Australasian origin, but has become more cosmopolitan. One branch of Eupetoidea contains three small families. These include the rail-babbler (Eupetidae) of Southeast Asia, the rockfowl (Picathartidae) of tropical Africa, and the rockjumpers (Chaetopidae) of Southern Africa.
The other branch is the family Petroicidae, the Australasian Robins. It contains roughly twice as many species than all of the other basal passerida. They can be found in Australia, New Guinea, New Zealand, the south Pacific to Fiji, Vanuatu, and the Lesser Sundas.
The main body of Passerida, which still contains over 1/3 of all bird species. It is divided into three large clades. I rank these as superfamilies: Sylvioidea, including the babblers and Old World warblers; Muscipapoidea, now expanded to include the waxwings, kinglets, and treecreepers, Old World flycatchers, thrushes, and starlings, and the Passeroidea, including weavers, finches, sparrows, buntings, New World warblers, cardinals, and tanagers. Oliveros et al. (2019) estimated that the split between Sylvioidea and the other clades happened around 26.9 mya and that the split between Muscipapoidea and Passeroidea occurred slightly later, about 26.6 mya.
Until recently it was unclear exactly which families were basal Passerida and which belonged in Corvida. Ericson et al. (2000) suggested that the nuclear c-myc could be used for this. They found an insertion in the gene of many passerine birds. None that were then thought to not be passerines had the insertion (this is still true). Other papers such as Ericson and Johansson (2003b), Jønsson and Fjeldså (2006a), Jønsson, Fjeldså, Ericson, and Irestedt (2007), and Irestedt and Ohlson (2008) pursued this idea further.
This idea successfully identified genera such as Chaetops, Eupetes, and Picathartes as members of Passerida. However, it excluded the most basal Passerida: Cnemophilidae (Satinbirds), Melanocharitidae (Berrypeckers), Notiomystidae (Stitchbird), and Callaeidae (New Zealand Wattlebirds).
it also failed when applied to the Australasian robins (Petroicidae), classifying them outside Passerida, even though it was successful with their closest cousins, Chaetops, Eupetes, and Picathartes. Exactly what caused this failure remains unclear. It may be a result of incomplete lineage sorting (ILS), once for the Eupetoidea, and once for the split between Petroicidae and the remaining Eupetoidea. The whole process from the split of Callaeoidea to the split of Petroicidae from the rest of Eupetoidea seems to have only taken about 1 1/2 million years, making the ILS hypothesis plausible.
Cnemophiloidea
The satinbirds (Cnemophilidae) used to be considered Corvida. In fact, they are the basal group in Passerida. These New Guinea endemics were once considered birds-of-paradise (Corvida: Paradisaeidae), but now have their own family within Passerida.
Cnemophilidae: Satinbirds Mayr, 1962
2 genera, 3 species Not HBW Family
- Yellow-breasted Satinbird / Silken Satinbird, Loboparadisea sericea
- Loria's Satinbird / Velvet Satinbird, Cnemophilus loriae
- Crested Satinbird / Antenna Satinbird, Cnemophilus macgregorii
Melanocharitoidea
Melanocharitidae: Berrypeckers & Longbills Sibley & Ahlquist, 1985
4 genera, 12 species HBW-13
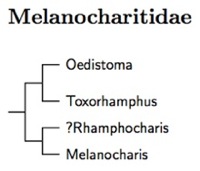 |
| Click for Melanocharitidae species tree |
|---|
The berrypeckers (Melanocharitidae) are another group of New Guinea endemics. Sibley and Monroe (1993) listed them in the Passeroidea, near the flowerpecker and sunbirds. They are now considered one of the basal groups in Passerida and follow the satinbirds in the linear order. Their placement here is based on the large scale studies by Oliveros et al. (2019), Kuhl et al. (2021), and Stiller et al. (2024). The berrypecker family includes the former honeyeaters Oedistoma and Toxorhamphus and the former flowerpeckers Melanocharis and Pristorhamphus.
The recently discovered Satin Berrypecker, Melanocharis citreola, was described by Milá et al. (2021).
The Spotted Berrypecker, Melanocharis crassirostris, has been split. The western subspecies crassirostris takes the new name Thick-billed Berrypecker, while the eastern subspecies piperata retains the English name Spotted Berrypecker. The two taxa differ in both morphology and plumage. The differences are more obvious in the females. See Beehler and Pratt (2016) and Gregory (2017).
The species tree is based on Milá et al. (2021). It contains a calibrated tree based on dates from Oliveros et al. (2019). Milá et al. found that the genus Rhamphocharis is embedded in Melanocharis. However, they also found that Melanocharis consists of two clades that separated roughly 9 million years ago. It makes sense to put them in separate genura. Fortunately, the name Pristorhamphus Finsch 1876 can be used for the smaller branch. The Fan-tailed Berrypecker, Pristorhamphus versteri is its type.
- Dwarf Longbill, Oedistoma iliolophus
- Pygmy Longbill, Oedistoma pygmaeum
- Yellow-bellied Longbill, Toxorhamphus novaeguineae
- Slaty-headed Longbill, Toxorhamphus poliopterus
- Fan-tailed Berrypecker, Pristorhamphus versteri
- Satin Berrypecker, Pristorhamphus citreolus
- Streaked Berrypecker, Pristorhamphus striativentris
- Obscure Berrypecker, Melanocharis arfakiana
- Black Berrypecker, Melanocharis nigra
- Mid-mountain Berrypecker, Melanocharis longicauda
- Thick-billed Berrypecker, Melanocharis crassirostris
- Spotted Berrypecker, Melanocharis piperata
Callaeoidea
The Callaeoidea include the stitchbird (Notiomystidae) and the New Zealand wattlebirds (Callaeidae). I follow Moyle et al. (2016), Oliveros et al. (2019), and Stiller et al. (2024) in placing them after the satinbirds (Cnemophilidae) and berrypeckers (Melanocharitidae).
Notiomystidae: Stitchbird Driskell et al., 2007
1 genus, 1 species HBW-14
The monotypic stitchbird was previously considered a honeyeater, but is actually most closely related to the wattlebirds (Ewen et al., 2006; Driskell et al., 2007). The split between the Stitchbird and the New Zealand wattlebirds happened about 26 mya (Oliveros et al., 2019). That and the obvious differences between them justfies considering it a monotypic family.
- Stitchbird / Hihi, Notiomystis cincta
Callaeidae: New Zealand Wattlebirds Sundevall, 1836 (1831)
3 genera, 5 species HBW-14
 |
Wattlebird phylogeny follows Lubbe et al. (2022), where the Huia is more closely related to the saddlebacks than the kokakos. They estimate that the kokakos diverged from the others around 10.9 mya, with the huia/saddleback split at about 8.4 mya.
Following OSNZ, IOC, and HBW-14, the Kokako, Callaeas cinereus, is split into North Island Kokako, Callaeas wilsoni, and South Island Kokako, Callaeas cinereus. See also Double and Murphy (2002). Further, the Saddleback, Philesturnus carunculatus is split into North Island Saddleback, Philesturnus rufusater, and South Island Saddleback, Philesturnus carunculatus. See also Parker et al. (2013). More recently, Lubbe et al. (2022) estimated that the two kokakos diverged around 2.7 mya, with the two saddlebacks diverging slightly earlier, around 2.6 mya.
- North Island Kokako, Callaeas wilsoni
- South Island Kokako, Callaeas cinereus
- Huia, Heteralocha acutirostris
- North Island Saddleback, Philesturnus rufusater
- South Island Saddleback, Philesturnus carunculatus
Eupetoidea
The Eupetoidea are the next group to branch off separately from the Passerida tree, and branch they did! Although the branches are small, they are widespread. The first division is between the Australasian Robins (Petroicidae) and the other three familes. Since there are a lot of robin species and few of the others, we put the others first. The remaining part includes rail-babbler (Eupetidae) from SE Asia and two sister African families, the rockfowl (Picathartidae) of tropical Africa, and the rockjumpers (Chaetopidae) of southern Africa.
 More recent large-scale studies, including Oliveros et al. (2019), Kuhl
et al. (2021), and Stiller et al. (2024) disagree. They put Petroicidae
together with Picathartes and Chaetops in the sister group
of the rest of the passerines. That suggests the focus on the
c-myc gene was mistaken. It also allows for other explanations of
how passerines got to Africa. E.g., Picathartes and
Chaetops could have been part of an earlier radiation into Africa.
More recent large-scale studies, including Oliveros et al. (2019), Kuhl
et al. (2021), and Stiller et al. (2024) disagree. They put Petroicidae
together with Picathartes and Chaetops in the sister group
of the rest of the passerines. That suggests the focus on the
c-myc gene was mistaken. It also allows for other explanations of
how passerines got to Africa. E.g., Picathartes and
Chaetops could have been part of an earlier radiation into Africa.
According to Oliveros et al. (2019), Eupetoidea separated from the remaining Passerida in the late Oligocene, about 29.8 mya. Soon thereafter, geologically speaking, the Australian Robins separated from the rest of Eupetoidea. This happened about 27.4 mya. The split of the SE Asian Rail-babbler family (Eupetidae) from the two African families happened about 22 mya. Finally, the rockfowl (Picathartidae) and rockjumpers (Chaetopidae) parted ways just under 20 mya. The last two families are quite different in both appearance and habits, as befits separate families.
The rockfowl and rockjumpers were considered Incertae sedis by Sibley and Monroe (1993) and for some time there had been discussion about whether they are best considered corvid relatives or part of Passerida. Several papers in the 2000's made the case for placing them in Passerida (Barker et al., 2004; Beresford et al., 2005; Cracraft et al., 2004; Irestedt and Ohlson, 2008). Then research by Jønsson et al. (2007) found that the Rail-babbler belonged in the same clade. Moyle et al. (2016) suggested the group originated roughly 23 mya. Recent large-scale studies found dates that are a bit older. Kuhl et al. (2012) give 28.3 mya, Oliveros et al. (2019) are at 28 mya, and Stiller et al. (2024) date the Eupetoidea to 25 mya.
Eupetidae: Rail-babbler Bonaparte, 1850
1 genus, 1 species HBW-12
- Rail-babbler, Eupetes macrocerus
Picathartidae: Rockfowl Lowe, 1938
1 genus, 2 species HBW-12
- White-necked Rockfowl, Picathartes gymnocephalus
- Gray-necked Rockfowl, Picathartes oreas
Chaetopidae: Rockjumpers Fjeldså et al. 2015
1 genus, 2 species Not HBW Family
- Cape Rockjumper, Chaetops frenatus
- Orange-breasted Rockjumper / Drakensberg Rockjumper, Chaetops aurantius
Petroicidae: Australasian Robins Mathews, 1919-20
16 genera, 51 species HBW-12
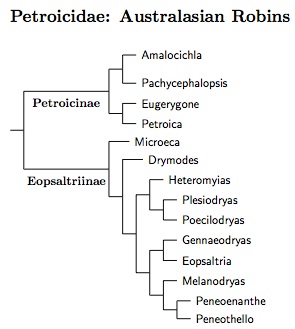 |
| Click for Petroicidae species tree |
|---|
For some time the arrangement of Petroicidae in TiF has been primarily based on Christidis et al. (2011). The papers by Loynes et al. (2009), and Miller and Lambert (2006) have also been consulted. The Christidis et al. paper is particularly useful as it includes over 80% of the Petroicidae recognized here.
Christidis et al. (2011) note six major clades in Petroicidae, and recommend that they be treated as subfamilies. I don't think this does a good job of describing the phylogeny, and prefer to separate Petroicidae into two subfamilies: Petroicinae and Eopsaltriinae. Oliveros et al. (2019) estimated the split between them happened around 24.4 mya. The two groups are similar enough that I rank them as subfamilies, not families.
One option would be to reduce the Christidis et al. (2011) subfamilies to tribes or less. If we ranked them as tribes, the subfamily Petroicinae would include monogeneric tribes Amalocichlini (Christidis et al., 2011) and Pachycephalopsini (Christidis et al., 2011), as well as Petroicini (including Eugerygone).
Possible tribes in Eopsaltriinae would be Drymodini (Wolters, 1980) and Microecini (Loynes et al., 2009), with the other species grouped in Eopsaltriini. There are no calibrated dates within either subfamily. Kearns et al. (2018, Fig. 3) and Kearns et al. (2019, Fig. 2) did estimate dates in Petroica, but the two papers disagree on the ages. My only takeway was that Petroica is likely best considered a single genus.
For the rest of the Petroicidae the only genetic evidence available to try to rank clade ages is information about ND2 differences. Unfortunately, Figure 1 of Christidis et al. (2011) makes it clear that the rate at which Petroicidae ND2 changes varies substantially. This means it is not particularly useful for determining genus boundaries.
English Name Changes
- The English name of Microeca griseoceps is changed from Yellow-legged Flycatcher / Yellow-legged Flyrobin to Yellow-legged Flyrobin.
- The English name of Microeca flavigaster is changed from Lemon-bellied Flycatcher / Lemon-bellied Flyrobin to Lemon-bellied Flyrobin.
Genus Changes
Genera in the current version are based on Birds of the World of May 2025. However, I've altered their treatment in two ways.
First, I don't think it makes sense to put the Black-chinned Robin, Poecilodryas brachyura, in genus Leucophantes. Based on archived sequence data, Laurent Raty found it is sister to Poecilodryas. As it appears rather similar to some species of Poecilodryas, I think it's more natural to include it in Poecilodryas rather than separate it in Leucophantes.
Second, I have also chosen to keep the distinctive Banded Yellow Robin in the monotypic genus Gennaeodryas Mathews 1920, rather than include it in Eopsaltria.
This implies the following genus changes:
- The Canary Flyrobin, Microeca papuana, is moved to the monotypic genus Devioeca Mathews 1925.
- The Olive Flyrobin, Microeca flavovirescens, and Yellow-legged Flyrobin, Microeca griseoceps, are placed genus Kempiella Mathews 1913, type K. griseoceps kempi.
- The Yellow-bellied Flyrobin, Microeca flaviventris, has been placed in the monotypic genus Cryptomicroeca Christidis, Irestedt, Rowe, Boles and Norman 2012.
- The Torrent Flyrobin, Microeca muelleriana, has been placed in the monotypic genus Monachella Savadori 1874.
- The genera Peneoenanthe and Peneothello have been absorbed by genus Melanodryas Gould 1865, type cucullata.
Species Splits
Based on Miller and Lambert (2006) and H&M-4 (among others), the New Zealand Robin, Petroica australis, has been split into North Island Robin, Petroica longipes, and South Island Robin, Petroica australis (inc. rakiura).
Based on Kearns et al. (2016), the Pacific Robin, Petroica multicolor, has been split into Pacific Robin, Petroica pusilla and the monotypic Norfolk Robin, Petroica multicolor.
Further, the Solomons Robin, Petroica polymorpha, including subspecies septentrionalis, kulambangrae, and dennisi, has been split from Pacific Robin, Petroica pusilla. See Kearns et al. (2016, 2018, 2019, 2020).
One of the New Guinea races of Drymodes, brevirostris, is no more closely related to superciliaris than to brunneopygia. I follow Christidis et al.'s suggestion to split it and the other New Guinea races as Papuan Scrub-Robin, Drymodes beccarii. They also suggest treating Melanodryas cucullata westralensis as a separate species as M. vittata otherwise renders M. cucullata paraphyletic. If this were a case of allopatric subspecies with distinct plumage or morphology, I would agree in a second. However, these are virtually indistinguishable taxa with ranges that abut one another across a good chunk of Australia. I suspect introgression here, and think that splitting them would be premature. Christidis et al. also note several other taxa in need of closer study.
The Black-capped Robin, Heteromyias armiti, including rothschildi, has been split from Ashy Robin, Heteromyias albispecularis, based on genetics, plumages, and calls. See Christidis et al. (2011), Beehler and Pratt (2016), and Gregory (2017).
Petroicidae Species List Mathews, 1919-20
Petroicinae Mathews, 1919-20
- Lesser Ground-Robin, Amalocichla incerta
- Greater Ground-Robin, Amalocichla sclateriana
- Green-backed Robin, Pachycephalopsis hattamensis
- White-eyed Robin, Pachycephalopsis poliosoma
- Garnet Robin, Eugerygone rubra
- Tomtit, Petroica macrocephala
- Black Robin, Petroica traversi
- North Island Robin, Petroica longipes
- South Island Robin, Petroica australis
- Scarlet Robin, Petroica boodang
- Flame Robin, Petroica phoenicea
- Rose Robin, Petroica rosea
- Pink Robin, Petroica rodinogaster
- Mountain Robin, Petroica bivittata
- Snow Mountains Robin, Petroica archboldi
- Norfolk Robin, Petroica multicolor
- Red-capped Robin, Petroica goodenovii
- Solomons Robin, Petroica polymorpha
- Pacific Robin, Petroica pusilla
Eopsaltriinae Mathews, 1946
- Canary Flyrobin, Devioeca papuana
- Olive Flyrobin, Kempiella flavovirescens
- Yellow-legged Flyrobin, Kempiella griseoceps
- Yellow-bellied Flyrobin, Cryptomicroeca flaviventris
- Torrent Flyrobin, Monachella muelleriana
- Jacky Winter, Microeca fascinans
- Lemon-bellied Flyrobin, Microeca flavigaster
- Golden-bellied Flyrobin, Microeca hemixantha
- Southern Scrub-Robin, Drymodes brunneopygia
- Papuan Scrub-Robin, Drymodes beccarii
- Northern Scrub-Robin, Drymodes superciliaris
- Gray-headed Robin, Heteromyias cinereifrons
- Ashy Robin, Heteromyias albispecularis
- Black-capped Robin, Heteromyias armiti
- Black-throated Robin, Plesiodryas albonotata
- Black-chinned Robin, Poecilodryas brachyura
- Black-sided Robin, Poecilodryas hypoleuca
- Buff-sided Robin, Poecilodryas cerviniventris
- White-browed Robin, Poecilodryas superciliosa
- Banded Yellow Robin, Gennaeodryas placens
- Western Yellow Robin, Eopsaltria griseogularis
- Eastern Yellow Robin, Eopsaltria australis
- White-breasted Robin, Eopsaltria georgiana
- White-faced Robin, Eopsaltria leucops
- Pale-yellow Robin, Eopsaltria capito
- Hooded Robin, Melanodryas cucullata
- Dusky Robin, Melanodryas vittata
- White-rumped Robin, Melanodryas bimaculata
- Mangrove Robin, Melanodryas pulverulenta
- White-winged Robin, Melanodryas sigillata
- Smoky Robin, Melanodryas cryptoleuca
- Slaty Robin, Melanodryas cyanus