The Babbler Clade
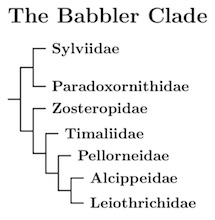 The babblers were long been treated as a wastebin taxon, with many
disparate species dumped in the group. They've sometimes been combined
with the Sylviidae, forming an even more disparate group. The previous
pages have covered many of the old world warbler families, leaving us
with a small sylviid core together with the babblers. The tree to the
right is based primarly on Cai et al. (2019), Oliveros et al. (2019), and
Stiller et al. (2024). It's similar to a previous such tree based on
Moyle et al. (2012), Fregin et al. (2012, Fig. 1 only), and Gelang et al.
(2009). I interpret these papers as evidence in favor of grouping the
remaining 450+ babblers, sylviids, and allies into 6 families.
The babblers were long been treated as a wastebin taxon, with many
disparate species dumped in the group. They've sometimes been combined
with the Sylviidae, forming an even more disparate group. The previous
pages have covered many of the old world warbler families, leaving us
with a small sylviid core together with the babblers. The tree to the
right is based primarly on Cai et al. (2019), Oliveros et al. (2019), and
Stiller et al. (2024). It's similar to a previous such tree based on
Moyle et al. (2012), Fregin et al. (2012, Fig. 1 only), and Gelang et al.
(2009). I interpret these papers as evidence in favor of grouping the
remaining 450+ babblers, sylviids, and allies into 6 families.
One of these families is the new Sylviidae, which has lost many of the old world warblers to new families such as Phylloscopidae and Acrocephalidae. It is sister to the parrotbills, Paradoxornithidae (once known as Panuridae, but Panurus is now on its own in a family of one, sister to the larks). The white-eyes remain in an expanded Zosteropidae. Then comes a more narrowly circumscribed Timaliidae, followed by the Pellorneidae and Leiothrichidae.
A natural name for Leiothrichidae is Turdoididae, which was sometimes used in the early 20th century. However, Leiothrix is the type genus of the family, so it is called Leiothrichidae.
As happens elsewhere in the Passerida, it doesn't make much sense to use roughly 20+ million years separation to define families. There has been a lot of recent differentiation, making this a very speciose section of the phylogeny. Because of that, using a 20 million year cutoff would make families would be large. To keep them a reasonable size, I only require that they be on relatively deep branches, and be coherent.
In fact, the range of age estimates shows that they lack sufficient precision to make the fine distinctions needed in this portion of the tree.
Let's start with point estimates from the big three. Oliveros et al. (2019) estimated that all of the six remaining families in Sylvioidea are less than 18.7 million years old, with the youngest five less than 16 million years old. In Kuhl et al. (2021), the numbers were shorter, 16.8 and 15.5 mya, respectively; in Stiller et al. (2024), they shorter still, 14.1 and 13.4 mya.
Worse, the age estimates are only approximate. The error bars for the ages of all six families range from about 16 to 20 million years in Oliveros et al., 8.5 to 22.5 in Kuhl et al., and 11 to 18 million years in Stiller et al.
Whichever numbers you prefer, a 20 million year rule for families would combine them all into a single family. That family would contain almost 500 species and unite such disparate birds as whitethroats, parrotbills, white-eyes, scimitar-babblers, and laughingthrushes. In fact, the 20 million rule combined with age estimates from Stiller et al. (2024) would include the larks and reedling too! That would create a family with over 1,280 species!
To make the families informative, we must allow them to be substantially younger than 20 million years. This problem will be even worse when we get consider Passeroidea. The solution is to not use a uniform temporal criterion for families across the avian tree. We allow the younger families in order to better organize the many species in Sylvioidea.
Sylviidae: Sylviid Warblers Leach, 1820
4 genera, 37 species HBW-11
In a sense, the Sylviidae are the epicenter of the DNA-driven taxonomic earthquake that has shaken up all of Passeriformes since the 1980's. At times, Sylviidae was considered a huge family, encompassing hundreds of Old World warblers. Now it has less than 40.
HBW-11 (del Hoyo et al., 2006), which uses a taxonomy current when the series started in 1992, lists 271 species in Sylviidae. In the TiF list, most of those have been spread across Acrocephalidae, Bernieridae, Cettiidae, Locustellidae, Macrosphenidae, and Phylloscopidae, with a few scattered elsewhere. And that was after HBW had already split off 145 species into Cisticolidae! At times, the babbler family Timaliidae, has also been considered a large family. In HBW-12 (del Hoyo et al., 2007) it contains 309 species.
That was then, this is now! As more genetic studies have been done, it has become clear that the core of Sylviidae has its true affinities with the babblers, not the other Old World warblers. Cibois (2003) even went so far as to suggest suppressing the name Sylviidae in favor of Timaliidae! Further analysis has clarified matters, and Sylviiade survives, but just barely. It has become a small family.
The changes in Sylvia itself mirror those of the Sylviidae. Most of the Sylvia warblers have been moved to Curruca, leaving only the Blackcap and Garden Warbler in Sylvia. In their place, Sylvia has been bolstered by former babblers: Horizorhinus (sometimes considered an Old World flycatcher) Lioptilus, Parophasma, and Pseudoalcippe, the latter species all from Timaliidae.
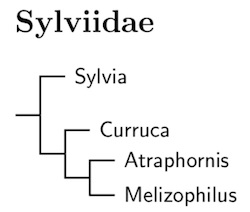 |
| Click for Sylviidae tree |
|---|
The arrangement of species here relies primarily on Cai et al. (2019).
It took a number of years to properly reconstruct the new Sylvia. Böhning-Gaese et al. (2003) presented evidence that the Blackcap and Garden Warbler are sister species, close to the Pseudoalcippe Hill-Babblers. Johansson et al. (2008b) found that the Bush Blackcap (Lioptilus) is closer to Pseudoalcippe than to Sylvia. Voelker et al. (2009) found a slightly different topology with Horizorhinus close to Pseudoalcippe. Although Voelker and Light (2011) did not include the Bush Blackcap, they comment on some unpublished work concerning it. Moyle et al. (2012) included all 4 genera. Finally, Cai et al. (2019) included all of current Sylvia in the same analysis. I use their results here.
Accordingly, I've submerged Horizorhinus (Dohrn's Thrush-Babbler), Parophasma (Abyssinian Catbird), Lioptilus (Bush Blackcap), and the Pseudoalcippe Hill-Babblers into Sylvia.
Since the type of Sylvia is atricapilla, the remaining Sylvia need a different name. Prior to seeing Cai et al. (2019), I revived the name Curruca for the whole group. However, after considering genetic distances, appearance, and available names, I now think a better solution is to use three genera: Atraphornis for the desert warblers, and Curruca and Melizophilus for the rest.
The subalpine warbler complex has been split into three (see Brambilla et al., 2008; Svensson 2013). They are Moltoni's Warbler, Curruca subalpina, (also known as moltonii), Western Subalpine Warbler, Curruca inornata, and Eastern Subalpine Warbler, Curruca cantillans. The name subalpina instead of moltonii seems to be preferred for Moltoni's Warbler, although the applicability of subalpina is not clear-cut. See Baccetti et al., (2007) and Svensson (2013).
Olsson et al. (2013) found that except for the already split minula, the “curruca” subspecies east of the Urals (and maybe some birds west of the Urals) are more closely related to Hume's Whitethroat, C. althaea than to curruca. There appear to be at least 4 species here. Although I'd prefer to have claification on things such as range boundaries, songs, and hybridization, I don't. This means that we have 2 choices, either group all 4 clades together as Hume's, or give treat them as species without knowning the whole story. I had originally done the first, but now think the second approach is better as it highlights the problem.
The lesser whitethroat complex includes:
- Desert Whitethroat, Curruca minula
- Lesser Whitethroat, Curruca curruca (inc. caucasica)
- *Stolzmann's Whitethroat, Curruca margelanica (inc. telengitica, chuancheica)
- *Steppe Whitethroat, Curruca halimodendri (inc. jaxartica)
- *Siberian Whitethroat, Curruca blythi
- Mountain Whitethroat / Hume's Whitethroat, Curruca althaea (inc. monticola)
where the asterisks indicate the new species. The English names posed something of a problem. Siberian for blythi was easy enough. The appellations Steppe and Central Asian have been used for halimodendri while althaea goes by Hume's and Mountain, with Mountain being more descriptive. The biggest problem is margelanica. The term Margelanic has been used, based on a city (Margilan) in eastern Uzbekistan. Unfortunately, it is doubly misleading. Margilan is not in the breeding range and Margelanic sounds like it belongs in S. America. Other suggestions include Mongolian or Gobi, but they don't seem quite right either. I thought about Chinese, but that equally applies to minula. The subspecies margelanica was described by Stolzmann, so lacking a better term, I've put his name on it.
- Eurasian Blackcap, Sylvia atricapilla
- Garden Warbler, Sylvia borin
- Dohrn's Thrush-babbler / Principe Flycatcher-Babbler, Sylvia dohrni
- Abyssinian Catbird, Sylvia galinieri
- Bush Blackcap, Sylvia nigricapillus
- Ruwenzori Hill-Babbler, Sylvia atriceps
- African Hill-Babbler, Sylvia abyssinica
- Barred Warbler, Curruca nisoria
- Layard's Warbler, Curruca layardi
- Banded Parisoma, Curruca boehmi
- Chestnut-vented Warbler, Curruca subcaerulea
- Yemen Warbler, Curruca buryi
- Brown Warbler / Brown Parisoma, Curruca lugens
- Arabian Warbler, Curruca leucomelaena
- Western Orphean-Warbler, Curruca hortensis
- Eastern Orphean-Warbler, Curruca crassirostris
- Desert Whitethroat, Curruca minula
- Lesser Whitethroat, Curruca curruca
- Stolzmann's Whitethroat, Curruca margelanica
- Steppe Whitethroat, Curruca halimodendri
- Siberian Whitethroat, Curruca blythi
- Mountain Whitethroat / Hume's Whitethroat, Curruca althaea
- African Desert-Warbler, Atraphornis deserti
- Asian Desert-Warbler, Atraphornis nana
- Tristram's Warbler, Melizophilus deserticola
- Menetries's Warbler, Melizophilus mystaceus
- Common Whitethroat, Melizophilus communis
- Spectacled Warbler, Melizophilus conspicillatus
- Marmora's Warbler, Melizophilus sardus
- Dartford Warbler, Melizophilus undatus
- Balearic Warbler, Melizophilus balearicus
- Cyprus Warbler, Melizophilus melanothorax
- Rueppell's Warbler, Melizophilus ruppeli
- Sardinian Warbler, Melizophilus melanocephalus
- Moltoni's Warbler, Melizophilus subalpina
- Western Subalpine Warbler, Melizophilus inornatus
- Eastern Subalpine Warbler, Melizophilus cantillans
Paradoxornithidae: Parrotbills, Fulvettas Horsfield & Moore, 1854
13 genera, 38 species HBW-12
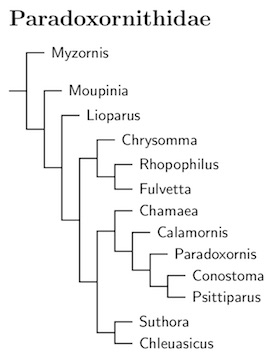 |
| Click for Paradoxornithidae tree |
|---|
The parrotbills form a small family that are sister to the sylviid warblers (Sylviidae). With one exception, the Wrentit of western North America, the parrotbills are native to Asia. Since the Sylviidae are primarily birds of Africa and the Palearctic, there is only limited geographic overlap between the two families.
The parrotbill family was previously called Panuridae. However, the genus Panurus (Bearded Reedling, aka Bearded Tit, Bearded Parrotbill) was found to be related to the larks, not to the other parrotbills. The name Paradoxornithidae is next in the priority line, and is applied here.
Unlike its sister clade Sylviidae, Paradoxornithidae has grown. It lost the reedling, but has added the Fulvetta fulvettas (formerly part of Alcippe, see Pasquet et al, 2006; Collar and Robson, 2007), another fulvetta (now Lioparus, formerly Alcippe), a former cisticolid (Rhopophilus), and five babblers: the two Chrysomma, the Wrentit (Chamaea), Moupinia and the highly distinctive Myzornis (Cibois, 2003; Gelang et al. 2009; Moyle et al., 2012).
The current arrangement of taxa is based on Cai et al. (2019). However, it is not much different from the previous one using Moyle et al. (2012), Pasquet et al. (2006), and Yeung et al. (2011).
Leader et al. (2013) found that the Chinese Hill Warbler, Rhopophilus pekinensis, consists of two species. Accordingly, it has been split into Tarim Babbler, Rhopophilus albosuperciliaris, and Beijing Babbler, Rhopophilus pekinensis.
Yeung et al. (2011) separately considered both nuclear and mitochondrial genes and found four main groups: the Fulvetta group, Suthora, Chleuasicus, and the Calamornis-Psittiparus group. Cai et al. (2019) modify this a bit to include Chamaea in the Calamornis-Psittiparus group (or Chamaea-Psittiparus group). I follow Cai et al. and make a couple of alterations to the Calamornis-Psittiparus group. The Reed Parrotbill, Paradoxornis heudei, has been placed in the monotypic genus Calamornis and Cholornis has been merged into Conostoma.
There seems to be insufficient reason to consider Northern Parrotbill, Calamornis polivanovi, as a separate species from Reed Parrotbill, Calamornis heudei, so they are merged as Reed Parrotbill, Calamornis heudei (see Penhallurick and Robson, 2009; Yeung et al., 2011).
The distinctive Yunnan Parrotbill, Chleuasicus ricketti, has been split from the Brown-winged Parrotbill, Chleuasicus brunneus. See Yeung et al. (2011) for genetic support and Wright et al. (2014) for photos.
- Fire-tailed Myzornis, Myzornis pyrrhoura
- Rufous-tailed Babbler, Moupinia poecilotis
- Golden-breasted Fulvetta, Lioparus chrysotis
- Yellow-eyed Babbler, Chrysomma sinense
- Jerdon's Babbler, Chrysomma altirostre
- Tarim Babbler, Rhopophilus albosuperciliaris
- Beijing Babbler, Rhopophilus pekinensis
- Spectacled Fulvetta, Fulvetta ruficapilla
- Indochinese Fulvetta, Fulvetta danisi
- Chinese Fulvetta, Fulvetta striaticollis
- White-browed Fulvetta, Fulvetta vinipectus
- Brown-throated Fulvetta, Fulvetta ludlowi
- Manipur Fulvetta, Fulvetta manipurensis
- Gray-hooded Fulvetta, Fulvetta cinereiceps
- Taiwan Fulvetta, Fulvetta formosana
- Wrentit, Chamaea fasciata
- Reed Parrotbill, Calamornis heudei
- Black-breasted Parrotbill, Paradoxornis flavirostris
- Spot-breasted Parrotbill, Paradoxornis guttaticollis
- Great Parrotbill, Conostoma aemodium
- Brown Parrotbill, Conostoma unicolor
- Three-toed Parrotbill, Conostoma paradoxum
- White-breasted Parrotbill, Psittiparus ruficeps
- Rufous-headed Parrotbill, Psittiparus bakeri
- Gray-headed Parrotbill, Psittiparus gularis
- Black-headed Parrotbill, Psittiparus margaritae
- Short-tailed Parrotbill, Suthora davidiana
- Fulvous Parrotbill, Suthora fulvifrons
- Black-throated Parrotbill, Suthora nipalensis
- Golden Parrotbill, Suthora verreauxi
- Pale-billed Parrotbill, Chleuasicus atrosuperciliaris
- Przevalski's Parrotbill, Chleuasicus przewalskii
- Spectacled Parrotbill, Chleuasicus conspicillatus
- Gray-hooded Parrotbill, Chleuasicus zappeyi
- Vinous-throated Parrotbill, Chleuasicus webbianus
- Ashy-throated Parrotbill, Chleuasicus alphonsianus
- Yunnan Parrotbill, Chleuasicus ricketti
- Brown-winged Parrotbill, Chleuasicus brunneus
Zosteropidae: White-eyes Bonaparte, 1853
12 genera, 151 species HBW-13
Zosterops itself is known as a “great speciator” for its propensity to invade new areas and diversify into new species and subspecies (Diamond et al., 1976; E. Mayr and Diamond, 2001). Zosterops and its relatives have colonized and re-colonized the same islands and mountain ranges. Sometimes an island has been colonized by a single species that diversifies, sometimes an island has been invaded by various groups of white-eyes. This makes it hard or even impossible to sensibly group Zosteropidae species on geographic grounds. You need DNA to sort them out. Although many of the clades make geographic sense, the colonization and re-colonization means the overall picture is hard to understand geographically unless you overlay geography on the phylogeny.
What is a White-eye?
What is a white-eye? Well, it is usually a small bird with a prominent white eye ring. Of course, this is a naive definition and some members of the white-eye family (Zosteropidae) don't have eye-rings, while certain other small birds with white eye-rings are not part of Zosteropidae. More seriously, there are two parts to the question. What families are the closest relatives of the white-eyes, and what genera and species are part of the white-eye family.
Traditionally, white-eyes were considered close to the honeyeaters
(Meliphagidae, now part of Corvida), flowerpeckers (Dicaeidae,
Passeroidea), and sunbirds (Nectariniidae, Passeroidea). In modern
taxonomies those are divergent families, with the honeyeaters nowhere
near the others.
Sibley and Ahlquist (1990) got white-eye taxonomy on the right track, correctly finding that the white-eyes are sylvioids. Cibois et al. (2003) went further, placing the Zosteropidae firmly in the Sylviidae/Timaliidae group. More recent analysis have confirmed this (e.g., Fregin et al., 2012; Oliveros et al., 2019; Kuhl et al., 2021; Stiller et al,, 2024).
The white-eyes are sister to the core babbler familes: Timaliidae, Pellorneidae, and Leiothrichidea. They separated from that group roughly 14—17 million years ago. Stiler et al. (2024) say 14 mya, Kuhl et al. (2021) estimate 15.5 mya, and Oliveros et al. (2019) found 16.9 mya. That makes them all relatively young families. I think calling the white-eyes and other babbler groups families is justified because the babblers are quite speciose, containing over 400 species. The case is particularly easy for the white-eyes as most are easily recognizable as different from the other babbler families. This illustrates how using several families helps organize these birds in a more coherent fashion, which is the purpose of bird families.
Our notion of what is and is not in the white-eye family has also changed. Cibois et al. (2003) noticed a close relationship between the genera Zosterops and Yuhina (except for Yuhina zantholeuca, now placed in the vireo genus Erpornis. In combination with Cibois et al. (2002), this close relationship implied Zosterops was also close to a number of species then considered Stachyris babblers. These have been moved to the Zosteropidae. More recent papers such as Cai et al. (2019), Oliveros et al. (2019), and Vinciguerra et al. (2022), have clarified these relationships by constructing more complete Zosteropidae phylogenies.
Cinnamon Ibon not a White-eye:
 There have also been some surprising losses from the Zosteropidae. Van
Balen (2008) listed the Cinnamon Ibon, Hypocryptadius cinnamomeus,
in a separate monotypic subfamily. However, he suggested it may not
belong with the white-eyes at all. More recently, Moyle et al. (2009a)
showed that the Ibon is not a white-eye, and indicated it may not even be
a sylvioid. The final piece of this saga was the paper by Fjeldså
et al. (2010). They found that the Cinnamon Ibon belongs to Passeroidea
rather then Sylvioidea. It is actually in the Old World Sparrow family,
Passeridae.
There have also been some surprising losses from the Zosteropidae. Van
Balen (2008) listed the Cinnamon Ibon, Hypocryptadius cinnamomeus,
in a separate monotypic subfamily. However, he suggested it may not
belong with the white-eyes at all. More recently, Moyle et al. (2009a)
showed that the Ibon is not a white-eye, and indicated it may not even be
a sylvioid. The final piece of this saga was the paper by Fjeldså
et al. (2010). They found that the Cinnamon Ibon belongs to Passeroidea
rather then Sylvioidea. It is actually in the Old World Sparrow family,
Passeridae.
Madanga not a White-eye either: More recently, another species has been removed from the Zosteropidae. Alström et al. (2015a) found that the Madanga, formerly Madanga ruficollis (previously Heleia on TiF), is not a white-eye either! It turns out that it is another member of Passeroidea. In fact, it is a pipit in the genus Anthus! As such, it takes the scientific name Anthus ruficollis. Its closest relative is the Alpine Pipit, Anthus gutturalis.
Basal Groups
Earlier work work by Zhang et al. (2007) and Moyle et al. (2009a) has clarified how the yuhinas and former Stachyris babblers relate to the white-eyes, with Cibois et al. (2002), Cibois (2003), Collar and Robson (2007), and Moyle et al. (2009a) providing valuable information.
The recent, very comprehensive papers by Oliveros et al. (2019) and Vinciguerra et al. (2022) found similar results. The various Yuhinas do not form a monophyletic group, but are a basal grade in Zosteropidae. They have been split into 3 genera, Parayuhina (monotypic), Staphida, and Yuhina. The various Yuhina groups are comprised mostly of mainland Asian species. However, the only one I have seen, Staphida everetti, occurs on the island of Borneo rather than on the mainland.
After the yuhinas, there is a side branch that contains the Philippine Sterrhoptilus babblers and 5 species divided into four genera. The use of separete genera is justified by the fact that each looks different, even though the genetic distance between them is not so large. Three of the genera are monotypic: Apalopteron, containing the Bonin White-eye; a Philippine species, the Flame-templed Babbler, genus Dasycrotapha; and Cleptornis, the Golden White-eye from the Northern Marianas. The two pygmy-babblers that are sometimes placed in Dasycrotapha have been left where they actually belong, in Sterrhoptilus.
The fourth genus is Rukia, with two species from the Caroline Islands. No DNA is available for either Rukia, but they may be closely related to Cleptornis. However, there used to be a third species in genus Rukia, the Olive-colored White-eye, now Zosterops oleagineus. DNA has firmly placed it in Zosterops. It was removed from Rukia when it was moved to Zosterops. But it is possible that the other two Rukia really are sister to it, and also belong within Zosterops. A third possible location for Rukia is within Heleia. Because of the uncertainty about Rukia, both are colored blue on the list.
The main branch continues with the now reunited Zosterornis Striped-Babblers (no more "Zosterornis"). Zosterornis is another Philippine group.
The next group includes nearly all the rest of the non-Zosterops white-eyes (a few have been merged into Zosterops). Since pipit Madanga has been ejected, these would include the genera Oculocincta, Lophozosterops, and Heleia. Mayr (1965) argued that most of them, perhaps all, should be merged into Heleia. Mees (1969), disagreed and seemed to have won the point. However, they and Megazosterops form a recent clade (4.5 million years old) in Vinciguerra et al. (2023), and do not seem so different as to require two genera, so I side with Mayr here and call them all Heleia.
The next branch begins with the Rufescent Darkeye, Tephrozosterops stalkeri, at about 3 mya. After that, the rest of the family, over 110 species, is in genus Zosterops.
Zosterops Overview
Now we're ready to consider Zosterops itself. Our version of Zosterops includes the small genera Chlorocharis, Speirops and Woodfordia. Zosterops is a very large genus and very young genus, perhaps less than 2.5 million years old. A single lineage turned into over 110 in that short period of time, averaging a new lineage every 23,000 years. No wonder the white-eyes are called “great speciators”!
The white-eyes propensity to speciate has also created a confused biogeographic picture. White-eyes have gone wild in invading new areas and forming new species. Sometimes, the same island has been invaded by more than one lineage in a short amount of time. Islands that seemed to be populated by two or more closely related white-eyes in a single invasion that then split are now known to have undergone two or more independent invasions. Nearby islands don't necessarily contain closely related white-eyes, even when they have been considered conspecific. The diversity of white-eyes can be quite bewildering.
Although not included in Vinciguerra et al.'s (2023) comprehensive analysis, the Sri Lanka White-eye, Zosterops ceylonensis, appears to be oldest branch of Zosterops. Both Lim et al. (2019) and O'Connell et al. (2019) show this, although the dating differs between them.
Until recently, we lacked a good, mostly complete phylogeny of the white-eyes. Earlier versions of TiF relied on Moyle et al. (2009a) and Cox (2013) come closest. Moyle et al. focuse on the eastern white-eyes, while Cox concentrated on the African white-eyes. Unfortunately, the basal portions of their phylogenetic trees were somewhat incongruent (e.g., where should the Silvereye Z. lateralis go?). Things started to change with the appearance of Cai et al. (2019). More recently, the UCE study by Oliveros et al. (2021) and the sequel by Vinciguerra et al. (2022) provided a solid and mostly complete phylogeny of the white-eyes.
The Sri Lanka White-eye, Zosterops ceylonensis appears to be basal. After that, the genus Zosterops divides into three clades. The first branch is an Indo-African clade. The remained is then divided into an Asiatic clade and an Australasiatic/Oceanic clade. This is clear in Gwee et al. (2020) and Vinciguerra et al. (2022).
East Indian Clade: The hypothesized East Indian clade includes the atriceps and chloris superspecies. Given that some white-eyes thought to be closely related have turned out to be not so close, it wouldn't be surprising to find that some of the species tenetatively placed in this clade don't belong here. Indeed, the Ashy-bellied White-eye, Zosterops citrinella, was thought to belong to the chloris superspecies, but Moyle et al. (2009a) placed it elsewhere. I've included some species of uncertain affinities as basal members. Some of them are thought to be close to the atriceps clade.
It makes biogeographic sense to have the East Indian clade in a relatively basal position. The closest relatives to Zosterops are Heleia and “Zosterornis”. Both are restricted to the Malay Archipelago, with “Zosterornis” in the Philippines and Heleia primarily in Indonesia.
The other two clades are more solid, although support for them is still weak. There is an Australasian/Oceanic clade found by Moyle et al. (2009a), and there is the African clade studied in detail by Cox (2013).
Zosterops, African Clade
The African clade includes a basal group (the Oriental clade), followed by a number of African clades. The topology for the African species is based on Warren et al. (2006), Melo et al. (2011), Cox (2013), and Cox et al. (2014). The African clades are marked with letters A-F as in Cox (2013).
Oriental White-eyes: This clade includes the Japanese White-eye, Zosterops japonicus, and probably the Lowland White-eye, Zosterops meyeni. It also includes the Oriental White-eye, Zosterops palpebrosus, and its close cousin, the Sri Lanka White-eye, Zosterops ceylonensis. The range of the Oriental White-eye stretches from China to Arabia, providing a link between the eastern and African white-eyes.
Ancient Indian Ocean White-eyes (A): The first African white-eyes are the Ancient Indian Ocean White-eyes, Z. semiflavus through Z. mauritianus. These white-eyes occur on Maruritius, La Réunion, Grande Comore, and formerly the Seychelles (semiflavus). Warren et al. (2006) discovered that these white-eyes formed a distinct group, and that they were not closely related to the Recent Indian Ocean White-eyes (clade E3). In other words, there were two separate invasions of the the islands in the western Indian Ocean. This clade stems from the initial invasion.
Warren et al. (2006), Melo et al. (2011), and Cox et al. (2014) find this group to be quite basal, I think too much so to make biogeographic sense. I have followed Cox (2013) on this where it is merely the most basal African clade. The time calibrated phylogeny in Cox et al. (2014) suggests that this “ancient” clade may be only 2 million years old. Still, it is ancient compared to the 1 million year ago origin of clade (E3), the Recent Indian Ocean white-eyes.
Socotra White-eye (B): This is an interesting case. Kirwan (2007) argued that the 4 white-breasted subspecies of abyssinicus are closely related. Indeed, he even argued that socotranus and abyssinicus are virtually indistinguishable. Cox's (2013) results upset that conclusion. However, Cox found that the Socotran socotranus ended up on a separate branch from the mainland socotranus. The mainland socotranus group with abyssinicus. As a result, I'm treating the Socotran socotranus as a separate species, Socotra White-eye, Zosterops socotranus.
Middle African White-eyes (F): The restructing of the Montane and African Yellow White-eye is on display in this clade, which ranges across the mid-section of Africa from Nigeria and the Gulf of Guinea to Pemba Island (near Zanzibar). It contains 3 pieces of lowland and montane white-eyes mixed together with a couple of Speirops and Pemba White-eye (sometimes included in African Yellow). We start with a pair of highland taxa that appear to be sister, the Mount Cameron Speirops, Zosterops melanocephalus, and the newly split Kilimanjaro White-eye, Zosterops eurycricotus (from Montane White-eye, Zosterops poliogastrus). Next up is the Albertine White-eye, Zosterops stuhlmanni. This is a rebranded version of the previously split Kivu White-eye, which has added stuhlmanni and toroensis. As stuhlmanni has priority, the scientific name had to change, and since the range is now larger, encompassing a big chunk of the Albertine rift area, it also got a new English name. It also contains the Forest White-eye, Zosterops stenocricotus, which has been split from African Yellow White-eye, Zosterops senegalensis (see also Melo et al, 2011).
Abyssinian White-eye (D): These species are followed by the Abyssinian White-eye, Zosterops abyssinicus, represented in Cox et al. (2014) by socotranus. Kirwan (2007) argues that the 4 white-breasted subspecies of abyssinicus are closely related. Indeed, he argues that socotranus and abyssinicus are virtually indistinguishable. The other two are arabs, which is slightly different, and omoensis, which is more easily distinguishable. Indeed, Cox (2013) groups the mainland white-breasted abyssinicus together. Cox (2013) found that the yellow-breasted subspecies flavilateralis and jubaensis (Pale White-eye, Z. flavilateralis of clade E1) are not closely related to mainland socotranus, and the entire white-breasted group.
Gulf of Guinea White-eyes (C): The Gulf of Guinea White-eyes include the other two former Speirops (Principe and Black-capped), along with the Principe White-eye, Zosterops ficedulinus, Annobon White-eye, Zosterops griseovirescens, and Sao Tome White-eye, Zosterops feae. Note that the last has been split from Z. ficedulinus. One result of all this is that the Gulf of Guinea white-eyes result from two separate expansions.
Southern African White-eyes (E2): The Southern African White-eyes include two additional splits: South Pare White-eye, Zosterops winifredae, from Z. poliogastrus, and Southern Yellow White-eye, Zosterops anderssoni, from Z. senegalensis. I had previously use the name Nyasa White-eye, Zosterops stierlingi, but the extensive taxon sampling in Cox (2013) has changed the picture. The species clade includes not just Zosterops stierlingi, but also tongensis and at least some portion of anderssoni (samples from Angola and Zambia). This likely means the type of anderssoni (from Nambia) is also included here, so use of the name anderssoni is justified. A sample of anderssoni from Malawi grouped with the Cape White-eye. The one sample of quanzae showed up on an earlier branch, together with a couple of samples from heinrichi. Oddly, samples of heinrichi from the same location ended up grouped with the Cape White-eye. I suspect that further work will clarify where these belong. Cox (2013) also sampled kasaicus, but did not include it in any of the trees. According to the text, the samples ended up somewhere in the winifredae-virens group.
East African Clade (E1): Cox (2013) and Cox et al. (2014) have two species from East Africa next, the yellow-breasted `abyssinicus', Pale White-eye, Z. flavilateralis (including jubaensis) and its sister taxon Z. poliogastrus mbuluensis, which I've promoted to species rank as Mbulu White-eye. The treatment mbuluensis as a species is also supported by Meimberg et al. (2016). Thus we maintain the traditional distinction between the highland (mbuluensis) and lowland (flavilateralis) white-eyes, but in a new form. Note that many of the highland forms in the range of flavilateralis are not so closely related to it, but that mbuluensis is.
Recent Indian Ocean Clade (E3): The arrangement of these species is based on Warren et al. (2006), who sampled more genes than Cox (2013). There are still some questions concerning species boundaries within the maderaspatanus group of Indian Ocean white-eyes. I've split Anjouan White-eye, Zosterops anjuanensis, from Malagasy White-eye, Zosterops maderaspatanus, on the grounds that it is more distantly related to Z. maderaspatanus than either Z. kirki or Z. mayottensis. It has historically been called “Lesser White-eye”. I've also split Moheli White-eye, Zosterops comorensis, from Mayotte White-eye, Zosterops mayottensis (or in two steps from Malagasy White-eye, Zosterops maderaspatanus). The three taxa (maderaspatanus, comorensis, mayottensis) seem about equally distant. For similar reasons, I've also separated Aldabra White-eye, Zosterops aldabrensis, from Kirk's White-eye, Zosterops kirki (or in two steps from Malagasy White-eye, Zosterops maderaspatanus).
As far as the allocation of subspecies within the maderaspatanus clade of Indian Ocean White-eyes is concerned, the extinct Seychelles Yellow White-eye, Z. semiflavus, often considered a subspecies of Z. mayottensis, is treated as a separate species because it is not even part of this group, instead belonging to the “ancient Indian Ocean White-eyes”. This leaves voeltzkowi and likely menaiensis as subspecies of Z. maderaspatanus.
| Current Division of Traditional White-eyes (compared to HBW vol. 13) | |
|---|---|
| The headings indicate species in HBW-13. Below them are the allocation of subspecies to the species on the TiF list. The subspecies with question marks indicate either a lack of genetic testing (menaiensis), or a subspecies that was tested but not placed in the tree (kasaicus). The subspecies in red appeared in two different groups in Cox (2013) (anderssoni, gerhardi, and heinrichi). Meimberg et al. (2016) also supports species status for kikuyuensis, mbuluensis, and silvanus. Although they claim support for species status for kulalensis, it is certainly not evident from their genetic analysis, with only a single sample closely allied to poliogastrus. | |
| Abyssinian White-eye, Zosterops abyssinicus | |
| Socotra White-eye Z. socotranus (A) | socotranus (Socotra) |
| Abyssinian White-eye Z. abyssinicus (D) | abyssinicus, arabs, socotranus (mainland), omoensis |
| Pale White-eye Z. flavilateralis (E1) | jubaensis, flavilateralis |
| Cape White-eye, Zosterops pallidus | |
| Orange River White-eye Z. pallidus (E2) | pallidus, sundevalli |
| Cape White-eye Z. virens (E2) | capensis, caniviridis, virens, atmorii, |
| African Yellow White-eye, Zosterops senegalensis | |
| Green White-eye Z. stuhlmanni (F) | toroensis, reichenowi, stuhlmanni |
| Forest White-eye Z. stenocricotus (F) | stenocricotus |
| Northern Yellow White-eye Z. senegalensis (F) | senegalensis, jacksoni, demeryi, gerhardi |
| Angolan Yellow White-eye Z. quanzae (E2) | kasaicus, heinrichi, quanzae |
| Southern Yellow White-eye Z. anderssoni (E2) | anderssoni, tongensis, stierlingi, |
| Montane White-eye, Zosterops poliogastrus | |
| Broad-ringed White-eye Z. eurycricotus (F) | eurycricotus |
| Kikuyu White-eye Z. kikuyuensis (F) | kikuyuensis |
| Montane White-eye Z. poliogastrus (F) | poliogastrus, kaffensis, kulalensis, |
| Mbulu White-eye Z. mbuluensis (E1) | mbuluensis |
| Taita White-eye Z. silvanus (E2) | silvanus |
| South Pare White-eye Z. winifredae (E2) | winifredae |
| Malagasy White-eye, Zosterops maderaspatanus | |
| Anjouan White-eye Z. anjuanensis (E3) | anjuanensis |
| Aldabra White-eye, Z. aldabrensis (E3) | aldabrensis |
| Kirk's White-eye Z. kirki (E3) | kirki |
| Malagasy White-eye Z. maderaspatanus (E3) | maderaspatanus, voeltzkowi, menaiensis |
| Moheli White-eye Z. comorensis (E3) | comorensis |
| Letters in parentheses refer to clades in the tree. | |
Zosterops, Australasian/Oceanic Clade
The Australasian/Oceanic Clade includes three subclades: a western Australasian clade, an eastern Australasian clade, and a Micronesian/Melanesian clade.
East Asian/Oceanic Clade: This clade includes white-eyes from China, the Malay Archipelago, and western Micronesia (Rota and Yap). The second subclade contains the Yellowish White-eye, Zosterops nigrorum. This brings us to the last portion of Zosterops, the 3-part clade. The first subclade may actually be basal in Zosterops, with all other clades being subclades. However, Moyle et al. (2009a) prefer a different topology which is followed here. They found that the Mountain Blackeye, Chlorocharis emiliae, is actually a Zosterops, apparently closely related to the Mountain White-eye. The placement of the Rota White-eye, Zosterops rotensis, and the Plain (formerly Yap) White-eye, Zosterops hypolais is inspired by Slikas et al. (2000).
West Australasian Clade: I've split one species in the western clade. The two representatives of Z. palpebrosus were widely separated, one in this clade, one in clade II. I have promoted the one here (Z. palpebrosus melanurus) to species status. Although it has apparently been considered a separate species in the past (HBW13), I couldn't find an English name. I'm temporarily designating it “Sunda White-eye” on geographic grounds. When considered a separate species, melanurus has contained two subspecies: melanurus and unicus. The subspecies unicus is what Moyle et al. actually sampled. As buxtoni is known to hybridize with melanurus, and palpebrosus is rather distant genetically, it is provisionally treated as a subspecies of melanurus, even though it is visually distinct. The other races of Oriental White-eye remain with palpebrosus. This means the highland race in Sumatra (buxtoni) is in the melanurus group, while the lowland race (auriventer) belongs to palpebrosus.
This clade also include the Ashy-bellied White-eye, Zosterops citrinella, which had been considered to possibly belong to the chloris superspecies. It doesn't, unless the results of Moyle et al. (2009a) are highly misleading.
East Australasian Clade: The second subclade includes the griseotinctus superspecies of the Solomons and nearby islands (minus the Banded White-eye, Zosterops vellalavella, which does not really belong with the others) and the Lifou White-eyes, which may be sister species (Cornetti et al., 2015). It also contains the Silvereye, Zosterops lateralis, and closely related species. However, even without vellalavella, the rest of the griseotinctus group is still not monophyletic, with some closer to the Silvereye than to other supposed group members.
The Dark-eyed White-eye, Zosterops tetiparius, including paradoxus (sometimes called rendovae) has been split from the Solomons White-eye, Zosterops kulambangrae, based on H&M-4, IOC 6.2, and Moyle et al. (2009).
The rendovae problem
There is a long-running controversy over the name Zosterops rendovae. The origin of the problem dates back to the 1880's. Ramsay (1881) named a new species of white-eye from San Cristobal (Makira) Island as Tephras olivaceus. Tristram (1882) moved the species to Zosterops. This meant that it needed a new name as there already was a Zosterops olivaceus. Tristram named it rendovae and included both a statement that it applied to Ramsay's bird and a description. Unfortunately, the description was not a description of Ramsay's bird, but of a white-eye from Rendova Island, not San Cristobal. In other words, Tristram had unwittingly applied the name to two taxa at once. Later that same year, Ramsay (1882) provided another replacement name (ugiensis) for T. olivaceus. Still later that same year, Salvadori recognized the double definition by Tristram. He restricted the name rendovae to the Rendova island white-eyes and proposed another replacement, ramsayi, for T. olivaceus. As it happens, Z. ramsayi was both preoccupied (by Z. ramsayi Masters 1876) and a junior synonym of the existing names. Some confusion about the San Cristobal specimen led to two more names being applied to the white-eyes there (salomonensis and alberti), but they need not concern us further.
Salvadori's designation of the Rendova white-eyes as rendovae was generally accepted and until 1955, although there was some confusion about how many white-eyes were on San Cristobal (and which name to use). Eventually it became clear there was only one and that ugiensis was the oldest available name. That would make the Solomons White-eye, Zosterops rendovae rather than Zosterops kulambangrae as it includes the Rendova population.
Mees (1955) argued that Salvadori's description was irrelevant for a replacement name, and that rendovae properly applied to T. olivaceus. He coined a new name (paradoxus) for the Rendova population. In this case, the Gray-throated White-eye would be Zosterops rendovae instead of Zosterops ugiensis. This solution has also been adopted by some. Others, such as Galbraith (1957), have insisted that Salvadori is correctly regarded as first reviser. This conflict has created considerable confusion concerning rendovae.
There had been a recent trend to suppress rendovae, although I do not believe that it has been made official. Clements does not use it. Recently, both H&M-4 and IOC6.2 have brought it back. In doing so, they have illustrated the problem with using rendovae. For H&M-4, Z. rendovae is the Dark-eyed White-eye, consisting of subspecies rendovae = paradoxus and tetiparius. For IOC 6.2, Z. rendovae is the Gray-throated White-eye, with subspecies rendovae = ugiensis, oblitus, and hamlini. In other words, H&M-4 and IOC 6.2 have provided the perfect illustration of the rendovae problem.
Although I think H&M-4 are technically correct, regarding Salvadori as first reviser, I nonetheless continue to treat rendovae as supressed. Using it is an impediment to accurate communication. Thus I call the Rendova white-eyes Zosterops tetiparius paradoxus and the San Cristobal white-eyes Zosterops ugiensis ugiensis. The Dark-eyed White-eye is then Z. tetiparius and the Gray-throated White-eye is Z. ugiensis.
NB: The Biodiversity Heritage Library provides access to the relevant original papers.
Micronesian/Melanesian Clade: Moyle et al. also included two subspecies of Z. ugiensis (ugiensis and hamlini) in their analysis. They ended up in different parts of the Micronesian/Melanesian subclade, with ugiensis itself more closely related to the Micronesian branch. There does not appear to be an English name for Z. hamlini. Since the hamlini subspecies is restricted to Bougainville Island, I'm referring to it as the “Bougainville White-eye”. The other subspecies, Z. ugiensis oblitus, is restricted to Guadalcanal. Given the proximity of Guadalcanal to San Cristobal (home of Z. u. ugiensis, I'm treating it a subspecies of Z. ugiensis for the present.
It's not clear exactly where to put the Bridled White-eye, Zosterops conspicillatus. Slikas et al. (2000) argued that it was not conspecific with Zosterops semperi. However, some trees put it in a clade with Z. semperi and the Olive-colored White-eye, Zosterops oleagineus (formerly in Rukia). Moyle et al. (2009a) included Z. cinereus in a clade with those two and Z. ugiensis, but didn't analyze Z. conspicillatus. I've bundled them all together with the other member of the cinereus superspecies, Z. finschii.
Based on Hayes et al. (2016), the Gray-brown White-eye, Zosterops cinereus, has been split into Kosrae White-eye, Zosterops cinereus, and Gray-brown White-eye, Zosterops ponapensis.
The newly recognized Vanikoro White-eye, Zosterops gibbsi, (Dutson, 2008) is thought to be close to sanctaecrucis, which is probably near metcalfii and stressmani. Together, they are the other branch of the subclade.
Zosteropidae: White-eyes
12 genera, 151 species
- White-collared Yuhina, Parayuhina diademata
Click for Zosteropidae tree - Striated Yuhina, Staphida castaniceps
- Indochinese Yuhina, Staphida torqueola
- Black-chinned Yuhina, Yuhina nigrimenta
- Taiwan Yuhina, Yuhina brunneiceps
- Stripe-throated Yuhina, Yuhina gularis
- Rufous-vented Yuhina, Yuhina occipitalis
- White-naped Yuhina, Yuhina bakeri
- Whiskered Yuhina, Yuhina flavicollis
- Burmese Yuhina, Yuhina humilis
- Bonin White-eye, Apalopteron familiare
- Flame-templed Babbler, Dasycrotapha speciosa
- Golden White-eye, Cleptornis marchei
- Teardrop White-eye, Rukia ruki
- Long-billed White-eye, Rukia longirostra
- Visayan Pygmy-Babbler, Sterrhoptilus pygmaeus
- Mindanao Pygmy-Babbler, Sterrhoptilus plateni
- Rusty-crowned Babbler, Sterrhoptilus capitalis
- Golden-crowned Babbler, Sterrhoptilus dennistouni
- Calabarzon Babbler, Sterrhoptilus affinis
- Visayan Babbler, Sterrhoptilus nigrocapitatus
- Palawan Striped-Babbler, Zosterornis hypogrammicus
- Chestnut-faced Babbler, Zosterornis whiteheadi
- Luzon Striped-Babbler, Zosterornis striatus
- Panay Striped-Babbler, Zosterornis latistriatus
- Negros Striped-Babbler, Zosterornis nigrorum
- Mees's White-eye, Heleia javanica
- Gray-hooded White-eye, Heleia pinaiae
- Pygmy White-eye, Heleia squamifrons
- Black-masked White-eye / Mindanao White-eye, Heleia goodfellowi
- Streak-headed White-eye, Heleia squamiceps
- Yellow-ringed White-eye, Heleia wallacei
- Giant White-eye, Heleia palauensis
- Cream-browed White-eye, Heleia superciliaris
- Crested White-eye, Heleia dohertyi
- Thick-billed Heleia, Heleia crassirostris
- Spot-breasted Heleia, Heleia muelleri
- Rufescent Darkeye, Tephrozosterops stalkeri
- Sri Lanka White-eye, Zosterops ceylonensis
- Reunion Olive White-eye, Zosterops olivaceus
- Mauritius Olive White-eye, Zosterops chloronothos
- Karthala White-eye, Zosterops mouroniensis
- Reunion Gray White-eye, Zosterops borbonicus
- Mauritius Gray White-eye, Zosterops mauritianus
- Indian White-eye, Zosterops palpebrosus
- †Marianne White-eye, Zosterops semiflavus
- Green White-eye, Zosterops stuhlmanni
- Fernando Po Speirops, Zosterops brunneus
- Forest White-eye, Zosterops stenocricotus
- Kikuyu White-eye, Zosterops kikuyuensis
- Northern Yellow White-eye, Zosterops senegalensis
- Heuglin's White-eye, Zosterops poliogastrus
- Mount Cameroon Speirops, Zosterops melanocephalus
- Broad-ringed White-eye, Zosterops eurycricotus
- Principe White-eye, Zosterops ficedulinus
- Sao Tome White-eye, Zosterops feae
- Annobon White-eye, Zosterops griseovirescens
- Principe Speirops, Zosterops leucophaeus
- Black-capped Speirops, Zosterops lugubris
- Pale White-eye, Zosterops flavilateralis
- Mbulu White-eye, Zosterops mbuluensis
- Seychelles White-eye, Zosterops modestus
- Malagasy White-eye, Zosterops maderaspatanus
- Anjouan White-eye, Zosterops anjuanensis
- Kirk's White-eye, Zosterops kirki
- Aldabra White-eye, Zosterops aldabrensis
- Moheli White-eye, Zosterops comorensis
- Mayotte White-eye, Zosterops mayottensis
- Pemba White-eye, Zosterops vaughani
- Taita White-eye, Zosterops silvanus
- Angolan Yellow White-eye, Zosterops quanzae
- South Pare White-eye, Zosterops winifredae
- Southern Yellow White-eye, Zosterops anderssoni
- Cape White-eye, Zosterops virens
- Orange River White-eye, Zosterops pallidus
- Citrine White-eye, Zosterops semperi
- Yellowish White-eye, Zosterops nigrorum
- Everett's White-eye, Zosterops everetti
- Mountain Blackeye, Zosterops emiliae
- Small Lifou White-eye, Zosterops minutus
- Swinhoe's White-eye, Zosterops simplex
- Warbling White-eye, Zosterops japonicus
- Lowland White-eye, Zosterops meyeni
- Black-capped White-eye, Zosterops atricapilla
- Hume's White-eye, Zosterops auriventer
- Abyssinian White-eye, Zosterops abyssinicus
- Socotra White-eye, Zosterops socotranus
- Chestnut-flanked White-eye, Zosterops erythropleurus
- Plain White-eye, Zosterops hypolais
- Christmas White-eye, Zosterops natalis
- Javan White-eye, Zosterops flavus
- Meratus White-eye, Zosterops meratusensis
- Black-crowned White-eye, Zosterops atrifrons
- Black-ringed White-eye, Zosterops anomalus
- Pale-bellied White-eye, Zosterops consobrinorum
- Lemon-bellied White-eye, Zosterops chloris
- Wakatobi White-eye, Zosterops flavissimus
- Sangkar White-eye, Zosterops melanurus
- Canary White-eye, Zosterops luteus
- Buru White-eye, Zosterops buruensis
- Seram White-eye, Zosterops stalkeri
- Tagula White-eye, Zosterops meeki
- Black-fronted White-eye, Zosterops chrysolaemus
- Green-fronted White-eye, Zosterops minor
- Togian White-eye, Zosterops somadikartai
- Sangihe White-eye, Zosterops nehrkorni
- Ashy-bellied White-eye, Zosterops citrinella
- Morotai White-eye, Zosterops dehaani
- Cream-throated White-eye, Zosterops atriceps
- Bridled White-eye, Zosterops conspicillatus
- Olive-colored White-eye, Zosterops oleagineus
- Dusky White-eye, Zosterops finschii
- Gray-brown White-eye, Zosterops ponapensis
- Kosrae White-eye, Zosterops cinereus
- Bismarck White-eye, Zosterops hypoxanthus
- Vanikoro White-eye, Zosterops gibbsi
- Green-backed White-eye, Zosterops xanthochroa
- Capped White-eye, Zosterops fuscicapilla
- Oya Tabu White-eye, Zosterops crookshanki
- Ambon White-eye, Zosterops kuehni
- Biak White-eye, Zosterops mysorensis
- Papuan White-eye, Zosterops novaeguineae
- †White-chested White-eye, Zosterops albogularis
- Silvereye, Zosterops lateralis
- Golden-bellied White-eye, Zosterops uropygialis
- Pearl-bellied White-eye, Zosterops grayi
- Santa Cruz White-eye, Zosterops sanctaecrucis
- Sanford's White-eye, Zosterops lacertosus
- Bare-eyed White-eye, Zosterops superciliosus
- Vanuatu White-eye, Zosterops flavifrons
- Fiji White-eye, Zosterops explorator
- Samoan White-eye, Zosterops samoensis
- Slender-billed White-eye, Zosterops tenuirostris
- Rennell White-eye, Zosterops rennellianus
- Large Lifou White-eye, Zosterops inornatus
- †Robust White-eye, Zosterops strenuus
- Wangi-wangi White-eye, Zosterops paruhbesar
- Kolombangara White-eye, Zosterops murphyi
- Louisiade White-eye, Zosterops griseotinctus
- Rota White-eye, Zosterops rotensis
- Gray-throated White-eye, Zosterops ugiensis
- Malaita White-eye, Zosterops stresemanni
- Bougainville White-eye, Zosterops hamlini
- Yellow-throated White-eye, Zosterops metcalfii
- Guadalcanal White-eye, Zosterops oblitus
- Vella Lavella White-eye, Zosterops vellalavella
- Gizo White-eye, Zosterops luteirostris
- Solomons White-eye, Zosterops kulambangrae
- Ranongga White-eye, Zosterops splendidus
- Dark-eyed White-eye, Zosterops tetiparius
Timaliidae: Babblers, Scimitar-Babblers Bonaparte, 1840
11 genera, 54 species HBW-12
The name Timaliidae is often attributed to Vigors & Horsfield, 1827 (e.g. Bock, 1994), but this is incorrect (Olson, 1995). It is attributed to Bonaparte by Brodkorb (1978).
Kuhl et al. (2021) date the split of Timaliidae from Pellorneidae and Leiothrichidae at 14 mya. Oliveros et al. (2019) estimated its age at 16.9 mya. Stiller et al. (2024) put the split closer to the present, around 11.9 mya.
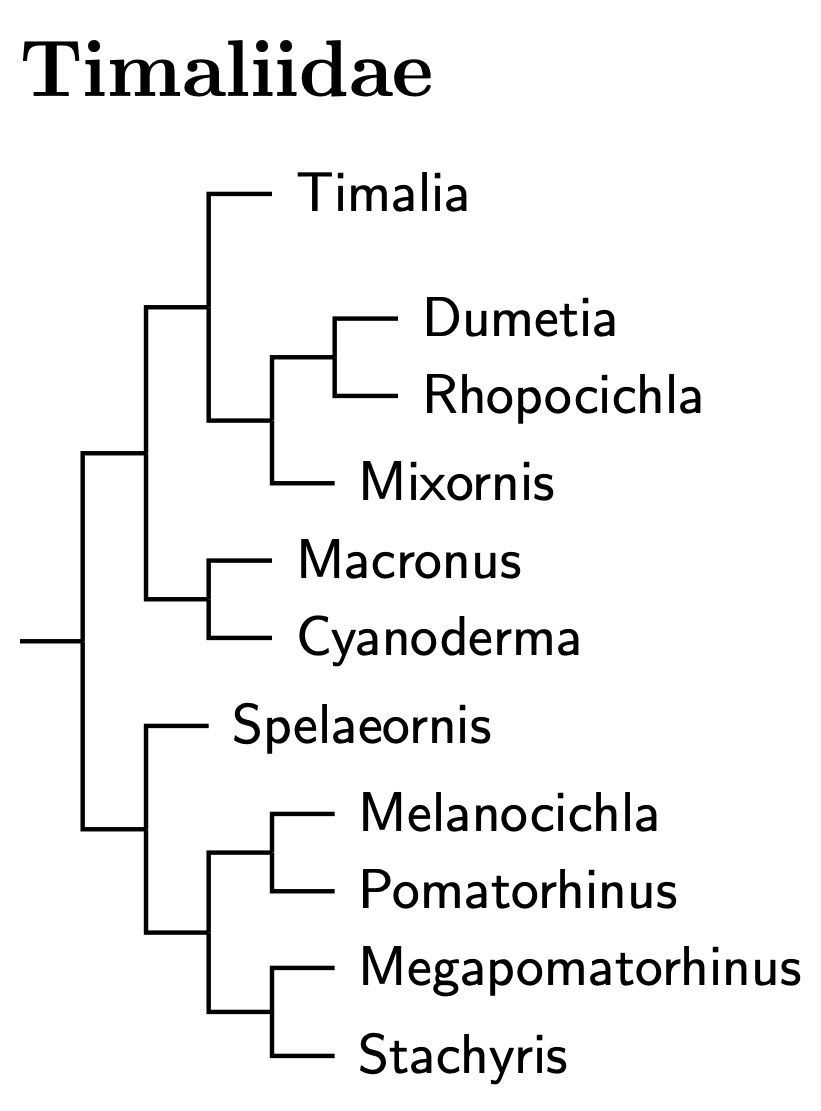 |
| Click for Timaliidae tree |
|---|
As with the other families in the babbler clade, the current arrangement is primarily based on Cai et al. (2019). In particular, this means that the basal clade consists of Timalia, which is sister to the Dumetia-Rhopocichla pair combined with Mixornis. I have left Rhopocichla and Dumetia in separate genera. Cai et al. (2020) puts their split around 6.5 mya, which neither compels nor precludes lumping the genera. I suspect the split is actually a bit more recent, but am not sure by how much.
Moyle et al. also found that the Pin-striped Tit-Babbler is more closely related to Timalia than to the remaining Macronus species. It is the type of Mixornis (Blyth, 1842). It seems likely that kelleyi, bornensis (split from gularis by Collar, 2006), and flavicollis are close relatives.
The other two Macronus are a bit more distant (Moyle et al., 2012), and sister to Cyanoderma. As for Cyanoderma, I had earlier followed Collar and Robson (2007), splitting Stachyridopsis from Stachyris. However, Moyle et al. (2012) found that the Chestnut-winged Babbler (formerly Stachyris erythroptera). also belongs in this group. It is the type species of Cyanoderma (Salvadori, 1874). This takes precendence over Stachyridopsis (Oates, 1883), and I have changed the names accordingly.
The other half of the babbler tree leads off with the Spelaeornis wren-babblers. They are sister to a pair of groups. The first includes the Pomatorhinus scimitar-babblers and, surprisingly, the Melanocichla laughingthrushes!
Cai et al. (2019) found that the two Melanocichla laughingthrushes, previously unsequenced, were not laughingthrushes at all. Rather, they are sister to Pomatorhinus. I had previously flagged them as uncertainly placed, leaving them near Ianothocincla and Pterorhinus, but I thought they were still some sort of laughingthrush. I was right to suspect them, but had no clue how different they really were from the laughingthrushes.
Gelang et al. (2009) showed that Xiphirhynchus is embedded in Pomatorhinus, so it has been merged into Pomatorhinus here. More recently, Dong et al. (2010) showed that Pomatorhinus should be split into two genera.
I had thought that Erythrogenys could be used, but that name was never actually made available. People thought it was available, but it is based on the misreading of a comment by Hodgson 1836. The correct name for that scimitar-babbler genus is Megapomatorhinus, Moyle et al. (2012). It has type hypoleucos. Megapomatorhinus is sister to Stachyris, which is why a separate genus is necessary. Finally, Moyle et al. also found that Sphenocichla is embedded in Stachyris.
The scimitar-babblers (Pomatorhinus and Megapomatorhinus) have been the subject of several detailed studies, including Dong et al. (2010, 2014), Nyári and Reddy (2013), and Reddy et al. (2011). It is likely that some of the existing species will be split, but the best way to do that is not yet clear.
What is clear is that species limits need to be changed for the Streak-breasted Scimitar-Babbler, Pomatorhinus ruficollis, Chestnut-backed Scimitar-Babbler, Pomatorhinus montanus, and White-browed Scimitar-Babbler, Pomatorhinus schisticeps. For now, the White-browed Scimitar-Babbler (P. schisticeps) loses all its subspecies that lie east of the Irrawaddy River. Three, nuchalis, fastidiosus, and klossi join the Chestnut-backed Scimitar-Babbler (P. montanus), while the olivaceus group (olivaceus, ripponi, difficilis, humilis, and annamensis) joins the Streak-breasted Scimitar-Babbler, P. ruficollis. Keep in mind that these are subject to further change.
- Chestnut-capped Babbler, Timalia pileata
- Tawny-bellied Babbler, Dumetia hyperythra
- Dark-fronted Babbler, Rhopocichla atriceps
- Pin-striped Tit-Babbler, Mixornis gularis
- Gray-cheeked Tit-Babbler, Mixornis flavicollis
- Gray-faced Tit-Babbler, Mixornis kelleyi
- Bold-striped Tit-Babbler, Mixornis bornensis
- Fluffy-backed Tit-Babbler, Macronus ptilosus
- Brown Tit-Babbler, Macronus striaticeps
- Golden Babbler, Cyanoderma chrysaeum
- Chestnut-winged Babbler, Cyanoderma erythropterum
- Crescent-chested Babbler, Cyanoderma melanothorax
- Rufous-fronted Babbler, Cyanoderma rufifrons
- Black-chinned Babbler, Cyanoderma pyrrhops
- Rufous-capped Babbler, Cyanoderma ruficeps
- Buff-chested Babbler, Cyanoderma ambiguum
- Bar-winged Wren-Babbler, Spelaeornis troglodytoides
- Rufous-throated Wren-Babbler, Spelaeornis caudatus
- Rusty-throated Wren-Babbler, Spelaeornis badeigularis
- Pale-throated Wren-Babbler, Spelaeornis kinneari
- Chin Hills Wren-Babbler, Spelaeornis oatesi
- Tawny-breasted Wren-Babbler, Spelaeornis longicaudatus
- Naga Wren-Babbler, Spelaeornis chocolatinus
- Gray-bellied Wren-Babbler, Spelaeornis reptatus
- Black Laughingthrush, Melanocichla lugubris
- Bare-headed Laughingthrush, Melanocichla calva
- Coral-billed Scimitar-Babbler, Pomatorhinus ferruginosus
- Red-billed Scimitar-Babbler, Pomatorhinus ochraceiceps
- Slender-billed Scimitar-Babbler, Pomatorhinus superciliaris
- Streak-breasted Scimitar-Babbler, Pomatorhinus ruficollis
- Taiwan Scimitar-Babbler, Pomatorhinus musicus
- White-browed Scimitar-Babbler, Pomatorhinus schisticeps
- Chestnut-backed Scimitar-Babbler, Pomatorhinus montanus
- Indian Scimitar-Babbler, Pomatorhinus horsfieldii
- Sri Lanka Scimitar-Babbler, Pomatorhinus melanurus
- Large Scimitar-Babbler, Megapomatorhinus hypoleucos
- Spot-breasted Scimitar-Babbler, Megapomatorhinus mcclellandi
- Rusty-cheeked Scimitar-Babbler, Megapomatorhinus erythrogenys
- Black-streaked Scimitar-Babbler, Megapomatorhinus gravivox
- Gray-sided Scimitar-Babbler, Megapomatorhinus swinhoei
- Black-necklaced Scimitar-Babbler, Megapomatorhinus erythrocnemis
- Black-throated Babbler, Stachyris nigricollis
- White-breasted Babbler, Stachyris grammiceps
- Chestnut-rumped Babbler, Stachyris maculata
- Gray-throated Babbler, Stachyris nigriceps
- Gray-headed Babbler, Stachyris poliocephala
- Sikkim Wedge-billed Babbler, Stachyris humei
- Cachar Wedge-billed Babbler, Stachyris roberti
- Nonggang Babbler, Stachyris nonggangensis
- Sooty Babbler, Stachyris herberti
- Snowy-throated Babbler, Stachyris oglei
- Spot-necked Babbler, Stachyris strialata
- White-necked Babbler, Stachyris leucotis
- White-bibbed Babbler, Stachyris thoracica
Pellorneidae: Ground Babblers Delacour, 1946
15 genera, 60 species Not HBW Family
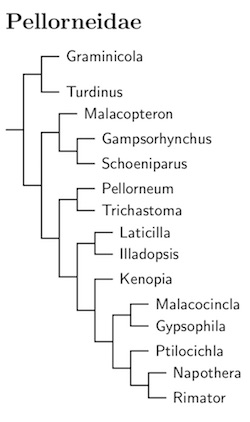 |
| Click for Pellorneidae tree |
|---|
I've characterized Pellorneidae as ground babblers because so many are ground and understory feeders.
The genus Alcippe is another poster child for the DNA revolution. The former Alcippe has been broken into four parts, with the Bush Blackcap ending up in Sylvia (Sylviidae) via Lioptilus, the Fulvetta fulvettas in Paradoxornithidae, the Schoeniparus fulvettas (including Pseudominla) here in Pellorneidae. The fate of the remaining Alcippe remained unclear until Cai et al. (2019, 2020) sampled all of them. They have joined Leiothrichidae as the subfamily Alcippeinae.
The most complete sampling of Pellorneidaeis Cai et al. (2019, 2020), and I have followed their arrangement of taxa whenever possible.
The basal taxa are Graminicola and Turdinus. All of the current Turdinus has previously been included in Napothera. Collar (2006) argued for separating them in Turdinus (see also Collar and Robson, 2007). As you can see on the tree, the birds in Turdinus have proven to be quite different from Napothera.
Working down the tree, Ophrydornis has been returned to Malacopteron. It is sister to Gampsorhynchus-Schoeniparus. Note that Schoeniparus, which was previously split from Alcippe (Pasquet et al., 2006; Collar and Robson, 2007), includes Pseudominla. The remaining portion of Pellorneidae is a well-supported clade in both Cai et al. (2019) and Moyle et al. (2012).
The basal branch is Pellorneum and Trichastoma. This version of Pellorneum has been reduced to three species, one of them uncertainly placed. The split between Pellorneum and Trichastoma is old enough that I have restored Trichastoma as a separate genus. It has absorbed a couple of species formerly in Malacocincla (malaccensis and cinereiceps). Cai et al. (2019) found that the Spot-throated Babbler, Trichastoma albiventre, belongs in this group. I'm guessing that the Marsh Babbler, Trichastoma palustre, belongs with it.
Olsson et al. (2013) found that the Rufous-vented Prinia, Laticilla burnesii does not belong in Prinia. It has been placed in Laticilla (Blyth 1845, type burnesii) and moved to the family Pellorneidae. The Swamp Prinia is thought to be a close relative, so it has become Laticilla cinerascens. The English names have been changed so they are grass-babblers rather than prinias..
Cai et al. (2019) sequenced L. burnesii and found it sister to Illadopsis. It's clear that the species in Illadopsis belong together (once Ptyrticus has been absorbed)
The remaining Pellorneidae form a clade sister to Laticilla--Illadopsis. The first branch is Kenopia. After it, there are two well-supported clades. One contains the remaining Malacocincla and the three Gypsophila (once part of Napothera). The other contains Ptilocichla, the one remaining Napothera species, and Rimator. Both Cai et al. and Moyle et al. recommend merging Rimator into Napothera, but I prefer to leave the distinctive Rimator group in its own genus. Note that Rimator includes Jabouilleia.
The genera Leonardina and Robsonius are no longer part of this family. Indeed, they are not even part of the larger babbler clade. Oliveros et al. (2012) found that Leonardina belongs to the Old World flycatcher family Muscicapidae and that Robsonius is in the grassbird family Locustellidae.
The Pale-breasted Thrush-Babbler, Illadopsis rufipennis may involve more than one species. Nguembock et al. (2009b) found samples of I. rufipennis extrema basal to a clade containing pyrrhoptera and other rufipennis.
The Rufous-rumped Grassbird, Graminicola bengalensis, has been split into Chinese Grassbird, Graminicola striatus, and Indian Grassbird, Graminicola bengalensis (Leader et al., 2010). As the English name suggests, the Chinese Grassbird includes the race sinicus.
Price et al. (2014) found L. burnesii closer to Napothera and Rimator than to Malacocincla. The primary English name of these two species has been changed from Prinia to Grass-Babbler (Rufous-vented Grass-Babbler and Swamp Grass-Babbler).
- Indian Grassbird, Graminicola bengalensis
- Chinese Grassbird, Graminicola striatus
- Marbled Wren-Babbler, Turdinus marmoratus
- Large Wren-Babbler, Turdinus macrodactylus
- Rusty-breasted Wren-Babbler, Turdinus rufipectus
- Black-throated Wren-Babbler, Turdinus atrigularis
- Sooty-capped Babbler, Malacopteron affine
- Gray-breasted Babbler, Malacopteron albogulare
- Scaly-crowned Babbler, Malacopteron cinereum
- Rufous-crowned Babbler, Malacopteron magnum
- Moustached Babbler, Malacopteron magnirostre
- Melodious Babbler, Malacopteron palawanense
- White-hooded Babbler, Gampsorhynchus rufulus
- Collared Babbler, Gampsorhynchus torquatus
- Rufous-throated Fulvetta, Schoeniparus rufogularis
- Rusty-capped Fulvetta, Schoeniparus dubius
- Dusky Fulvetta, Schoeniparus brunneus
- Yellow-throated Fulvetta, Schoeniparus cinereus
- Golden-fronted Fulvetta, Schoeniparus variegaticeps
- Rufous-winged Fulvetta, Schoeniparus castaneceps
- Black-crowned Fulvetta, Schoeniparus klossi
- Puff-throated Babbler, Pellorneum ruficeps
- Brown-capped Babbler, Pellorneum fuscocapillus
- Black-capped Babbler, Pellorneum capistratum
- Short-tailed Babbler, Trichastoma malaccensis
- Ashy-headed Babbler, Trichastoma cinereiceps
- Ferruginous Babbler, Trichastoma bicolor
- White-chested Babbler, Trichastoma rostratum
- Sulawesi Babbler, Trichastoma celebense
- Spot-throated Babbler, Trichastoma albiventre
- Marsh Babbler, Trichastoma palustre
- Temminck's Babbler, Trichastoma pyrrogenys
- Buff-breasted Babbler, Trichastoma tickelli
- Sumatran Babbler, Trichastoma buettikoferi
- Rufous-vented Grass-Babbler, Laticilla burnesii
- Swamp Grass-Babbler, Laticilla cinerascens
- Brown Illadopsis, Illadopsis fulvescens
- Pale-breasted Illadopsis, Illadopsis rufipennis
- Mountain Illadopsis, Illadopsis pyrrhoptera
- Blackcap Illadopsis, Illadopsis cleaveri
- Scaly-breasted Illadopsis, Illadopsis albipectus
- Spotted Thrush-Babbler, Illadopsis turdina
- Puvel's Illadopsis, Illadopsis puveli
- Rufous-winged Illadopsis, Illadopsis rufescens
- Striped Wren-Babbler, Kenopia striata
- Abbott's Babbler, Malacocincla abbotti
- Horsfield's Babbler, Malacocincla sepiaria
- Black-browed Babbler, Malacocincla perspicillata
- Mountain Wren-Babbler, Gypsophila crassa
- Streaked Wren-Babbler, Gypsophila brevicaudata
- Limestone Wren-Babbler, Gypsophila crispifrons
- Striated Wren-Babbler, Ptilocichla mindanensis
- Falcated Wren-Babbler, Ptilocichla falcata
- Bornean Wren-Babbler, Ptilocichla leucogrammica
- Eyebrowed Wren-Babbler, Napothera epilepidota
- Long-billed Wren-Babbler, Rimator malacoptilus
- White-throated Wren-Babbler, Rimator pasquieri
- Sumatran Wren-Babbler, Rimator albostriatus
- Naung Mung Scimitar-Babbler, Rimator naungmungensis
- Short-tailed Scimitar-Babbler, Rimator danjoui
Leiothrichidae: Laughingthrushes Swainson, 1831
23 genera, 143 species Not HBW Family
The final piece of the sylviid/babbler clade is Leiothrichidae, the laughingthrush family. As with the others, the current taxonomy follows Cai et al. (2019, 2020).
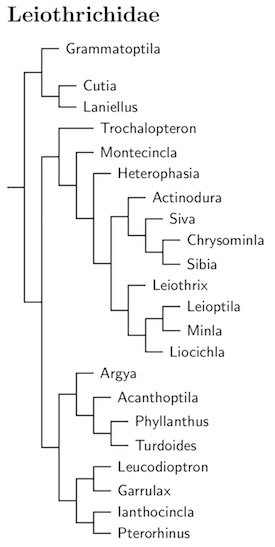 |
| Click for Leiothrichidae tree |
|---|
As you can see from the tree diagram, I've put Alcippe its own subfamily, Alcippeinae. Although some papers found it closer to the to the ground babbler family, Pellorneidae (Gelang et al., 2009; Cibois et al., 2018), support was low and they only sampled a single species of Alcippe. In contrast, papers that put Alcippeidae sister to the laughingthrush family, Leiothrichidae (Moyle et al., 2012; Cai et al., 2019, 2020) include some that sample all the Alcippeidae and have high support values.
Why a subfamily? Mainly because the division between Alcippeinae and Cai et al. (2020) found the split between Alcippeinae and Leiothrichinae is deep relative to the crown group of Leiothrichinae. Cai et al. (2020) also put origin of the two subfamilies around 17.5 mya. The big three disagree with this age. According to Oliveros et al. (2019), the root of Leiothrichidae is only 15 mya. Kuhl et al. (2021) put it at 12.1 mya, and Stiller et al. (2024) estimated it at only 11.9 mya. For the record, Cai et al. (2020) put that slightly earlier than the Alcippeinae-Leiothrichinae split, just under 18 mya. Obviously, these numbers don't match. Who's right here? I don't know.
Alcippeinae: Alcippe Fulvettas Cai et al., 2019
The current version of the genus Alcippe is another poster child for the DNA revolution. The former Alcippe has been broken into four parts, with the Bush Blackcap ending up in Sylvia (Sylviidae) via Lioptilus, the Fulvetta fulvettas in Paradoxornithidae, the Schoeniparus fulvettas (including Pseudominla) here in Pellorneidae, and the remaining Alcippe in Leiothrichidae.
As if that was not enough, the Gray-cheeked Fulvetta, Alcippe morrisonia has been split into several species based on Zou et al. (2007) and Song et al. (2009).
- Brown-cheeked Fulvetta, Alcippe poioicephala
- Black-browed Fulvetta, Alcippe grotei
- Brown Fulvetta, Alcippe brunneicauda
- Javan Fulvetta, Alcippe pyrrhoptera
- Nepal Fulvetta, Alcippe nipalensis
- David's Fulvetta, Alcippe davidi
- Yunnan Fulvetta, Alcippe fratercula
- Mountain Fulvetta, Alcippe peracensis
- Huet's Fulvetta, Alcippe hueti
- Gray-cheeked Fulvetta, Alcippe morrisonia
Leiothrichinae: Laughingthrushes Swainson, 1831
One little secret about the English name is that, like the Ground-Babblers, the Laughingthrushes also often feed on the groud.
The laughingthrush genus Garrulax itself has often been treated as a large genus. However, that doesn't work well as parts of it don't belong together. Before adequate genetic testing had been done, Collar and Robson broke Garrulax into 11 genera: Dryonastes, Melanocichla, Rhinocichla, Pterorhinus, Grammatoptila, Stactocichla, Leucodioptron, Strophocincla, Ianthocincla, and Trochalopteron, and Garrulax. They also grouped Babax with the laughingthrushes. We retain about half of these genera.
The monotypic Grammatoptila leads off a group in the basal position in Leiothrichidae, away from the other laughingthrushes. The other members of the group are Cutia and Laniellus. Following Gregory and Dickinson (2012), the genus name Laniellus Swainson, 1832 has replaced Crocias Temminck, 1836.
The family then divides in two. The first clade includes the Trochalopteron laughingthrushes and a number of smaller genera including the barwings, minlas, sibias, and other species. The first clade include Turdoides and a variety of Garrulax and former Garrulax laughingthrushes.
The first big piece of Leiothrichidae begins with the Trochalopteron laughingthrushes. Based on Robin et al. (2017), the Kerala Laughingthrush, Trochalopteron fairbanki, and Black-chinned Laughingthrush, Trochalopteron cachinnans, have been moved to Montecincla, one branch down from Trochalopteron. Further, there are splits:
- The Black-chinned Laughingthrush, Montecincla cachinnans, has been split into Nilgiri Laughingthrush, Montecincla cachinnans, and Banasura Laughingthrush, Montecincla jerdoni.
- The Kerala Laughingthrush, Trochalopteron fairbanki, has been split into Palani Laughingthrush, Montecincla fairbanki, and Ashambu Laughingthrush, Montecincla meridionalis.
After Montecincla are the Heterophasia sibias (including Malacias). The clade then splits into two subgroups.
The first subgroup includes the Sibia barwings, along with the two minlas (Siva and Chrysominla), and the Actinodura barwings. The second starts with Leiothrix (including Mesia), followed by the Minla-Leioptila pair, and Liocichla.
This brings us to the main laughingthrush clade, which also has two branches, the Turdoides group and the Garrulax group. The Turdoides group is sister to the main laughingthrush clade (containing Garrulax). Moyle et al. (2012) found that Phyllanthus and Kupeornis are embedded in it and this had previously led me to use a broad Turdoides. This was rearranged and divided taking Price et al. (2014) into account. The two embedded genera were restored, and Acanthoptila, Argya, Chatarrhaea, and Malacocircus were separated from Turdoides. This result ws similar to H&M-4 (Dickinson and Christidis, 2014) and the HBW BirdLife Checklist (del Hoyo and Collar, 2016), except that three species were placed in Malacocircus based on Price et al. (2014). Later, the two Malcomia species (malcolmi and cinereifrons) were separated using Cibois et al. (2018).
Cai et al. (2019), including their timeline, led to further changes. I submerged Malcolmia and Malacocircus into Argya. Further, Kupeornis was merged into Phyllanthus. The branching order here is now Argya, Acanthoptila, Phyllanthus, Turdoides.
The Garrulax group has also undergone various changes as we learned more about babbler taxonomy. Previously, I had divided the Garrulax group into six genera (one uncertainly placed): Leucodioptron, Stactocichla, Garrulax, Melanocichla, Ianthocincla, and Pterorhinus. Various papers gave various results for the arrangement of Pterorhinus. I based the structure of Pterorhinus on Päckert et al. (2011), with assistance from Luo et al. (2009), Moyle et al. (2012), Robin et al. (2017), and Wu et al. (2014).
That has all been swept away by Cai et al. (2019). Now there are four genera in two pairs: Leucodioptron, Garrulax, Ianthocincla, and Pterorhinus. One genus, Stactocichla, has been merged into Leucodioptron. The other missing genus, Melanocichla, has a more interesting fate. Cai et al. were the first to sequence it, and they found that the two Melanocichla are not laughingthrushes. They are not in Leiothrichidae at all, but belong in Timaliidae, sister to the Pomatorhinus scimitar-babblers!
- Striated Laughingthrush, Grammatoptila striata
- Himalayan Cutia, Cutia nipalensis
- Vietnamese Cutia, Cutia legalleni
- Gray-crowned Crocias, Laniellus langbianis
- Spotted Crocias, Laniellus albonotatus
- Scaly Laughingthrush, Trochalopteron subunicolor
- Brown-capped Laughingthrush, Trochalopteron austeni
- Blue-winged Laughingthrush, Trochalopteron squamatum
- Striped Laughingthrush, Trochalopteron virgatum
- Streaked Laughingthrush, Trochalopteron lineatum
- Bhutan Laughingthrush, Trochalopteron imbricatum
- Variegated Laughingthrush, Trochalopteron variegatum
- Black-faced Laughingthrush, Trochalopteron affine
- White-whiskered Laughingthrush, Trochalopteron morrisonianum
- Brown-cheeked Laughingthrush, Trochalopteron henrici
- Elliot's Laughingthrush, Trochalopteron elliotii
- Red-tailed Laughingthrush, Trochalopteron milnei
- Collared Laughingthrush, Trochalopteron yersini
- Chestnut-crowned Laughingthrush, Trochalopteron erythrocephalum
- Assam Laughingthrush, Trochalopteron chrysopterum
- Red-winged Laughingthrush, Trochalopteron formosum
- Malayan Laughingthrush, Trochalopteron peninsulae
- Silver-eared Laughingthrush, Trochalopteron melanostigma
- Golden-winged Laughingthrush, Trochalopteron ngoclinhense
- Banasura Laughingthrush, Montecincla jerdoni
- Nilgiri Laughingthrush, Montecincla cachinnans
- Palani Laughingthrush, Montecincla fairbanki
- Ashambu Laughingthrush, Montecincla meridionalis
- Long-tailed Sibia, Heterophasia picaoides
- White-eared Sibia, Heterophasia auricularis
- Rufous Sibia, Heterophasia capistrata
- Beautiful Sibia, Heterophasia pulchella
- Gray Sibia, Heterophasia gracilis
- Black-headed Sibia, Heterophasia desgodinsi
- Dark-backed Sibia, Heterophasia melanoleuca
- Hoary-throated Barwing, Sibia nipalensis
- Taiwan Barwing, Sibia morrisoniana
- Streak-throated Barwing, Sibia waldeni
- Streaked Barwing, Sibia souliei
- Blue-winged Minla, Siva cyanouroptera
- Bar-throated Minla, Chrysominla strigula
- Rusty-fronted Barwing, Actinodura egertoni
- Spectacled Barwing, Actinodura ramsayi
- Black-crowned Barwing, Actinodura sodangorum
- Red-billed Leiothrix, Leiothrix lutea
- Silver-eared Mesia, Leiothrix argentauris
- Red-tailed Minla, Minla ignotincta
- Rufous-backed Sibia, Leioptila annectens
- Bugun Liocichla, Liocichla bugunorum
- Emei Shan Liocichla, Liocichla omeiensis
- Steere's Liocichla, Liocichla steerii
- Red-faced Liocichla, Liocichla phoenicea
- Scarlet-faced Liocichla, Liocichla ripponi
- Large Gray Babbler, Argya malcolmi
- Ashy-headed Laughingthrush, Argya cinereifrons
- Rufous Babbler, Argya subrufa
- Slender-billed Babbler, Argya longirostris
- Orange-billed Babbler, Argya rufescens
- Jungle Babbler, Argya striata
- Yellow-billed Babbler, Argya affinis
- Rufous Chatterer, Argya rubiginosa
- Scaly Chatterer, Argya aylmeri
- Iraq Babbler, Argya altirostris
- Afghan Babbler, Argya huttoni
- Common Babbler, Argya caudata
- Fulvous Babbler, Argya fulva
- Arabian Babbler, Argya squamiceps
- Striated Babbler, Argya earlei
- White-throated Babbler, Argya gularis
- Spiny Babbler, Acanthoptila nipalensis
- Capuchin Babbler, Phyllanthus atripennis
- White-throated Mountain Babbler, Phyllanthus gilberti
- Chapin's Babbler, Phyllanthus chapini
- Red-collared Babbler, Phyllanthus rufocincta
- Brown Babbler, Turdoides plebejus
- White-rumped Babbler, Turdoides leucopygia
- Scaly Babbler, Turdoides squamulata
- White-headed Babbler, Turdoides leucocephala
- Arrow-marked Babbler, Turdoides jardineii
- Bare-cheeked Babbler, Turdoides gymnogenys
- Blackcap Babbler, Turdoides reinwardtii
- Dusky Babbler, Turdoides tenebrosa
- Southern Pied-Babbler, Turdoides bicolor
- Hartlaub's Babbler, Turdoides hartlaubii
- Black-lored Babbler, Turdoides sharpei
- Black-faced Babbler, Turdoides melanops
- Northern Pied-Babbler, Turdoides hypoleuca
- Hinde's Babbler, Turdoides hindei
- Chinese Hwamei, Leucodioptron canorum
- Taiwan Hwamei, Leucodioptron taewanum
- Spot-breasted Laughingthrush, Leucodioptron merulinum
- Orange-breasted Laughingthrush, Leucodioptron annamensis
- Lesser Necklaced Laughingthrush, Garrulax monileger
- Rufous-fronted Laughingthrush, Garrulax rufifrons
- Sunda Laughingthrush, Garrulax palliatus
- White-crested Laughingthrush, Garrulax leucolophus
- Sumatran Laughingthrush, Garrulax bicolor
- Black-hooded Laughingthrush, Garrulax milleti
- Cambodian Laughingthrush, Garrulax ferrarius
- White-necked Laughingthrush, Garrulax strepitans
- Gray Laughingthrush, Garrulax maesi
- Rufous-cheeked Laughingthrush, Garrulax castanotis
- Snowy-cheeked Laughingthrush, Ianthocincla sukatschewi
- Moustached Laughingthrush, Ianthocincla cineracea
- Rufous-chinned Laughingthrush, Ianthocincla rufogularis
- Chestnut-eared Laughingthrush, Ianthocincla konkakinhensis
- Spotted Laughingthrush, Ianthocincla ocellata
- Giant Laughingthrush, Ianthocincla maxima
- White-speckled Laughingthrush, Ianthocincla bieti
- Barred Laughingthrush, Ianthocincla lunulata
- Wynaad Laughingthrush, Pterorhinus delesserti
- Rufous-vented Laughingthrush, Pterorhinus gularis
- White-cheeked Laughingthrush, Pterorhinus vassali
- Yellow-throated Laughingthrush, Pterorhinus galbanus
- Blue-crowned Laughingthrush, Pterorhinus courtoisi
- Chestnut-capped Laughingthrush, Pterorhinus mitratus
- Chestnut-hooded Laughingthrush, Pterorhinus treacheri
- Rufous-necked Laughingthrush, Pterorhinus ruficollis
- Chestnut-backed Laughingthrush, Pterorhinus nuchalis
- Black-throated Laughingthrush, Pterorhinus chinensis
- White-browed Laughingthrush, Pterorhinus sannio
- Masked Laughingthrush, Pterorhinus perspicillatus
- Greater Necklaced Laughingthrush, Pterorhinus pectoralis
- Plain Laughingthrush, Pterorhinus davidi
- Mount Victoria Babax, Pterorhinus woodi
- Chinese Babax, Pterorhinus lanceolatus
- Giant Babax, Pterorhinus waddelli
- Tibetan Babax, Pterorhinus koslowi
- White-throated Laughingthrush, Pterorhinus albogularis
- Rufous-crowned Laughingthrush, Pterorhinus ruficeps
- Gray-sided Laughingthrush, Pterorhinus caerulatus
- Buffy Laughingthrush, Pterorhinus berthemyi
- Rusty Laughingthrush, Pterorhinus poecilorhynchus