Myrmotheridae: Antpittas MacGillivray, 1839
The family name for the Antpittas has been changed from Grallariidae to Myrmotheridae because the latter has priority. See Gaudin et al. (2021a).
Notes — Myrmotheridae
 |
| Click for Myrmotheridae species |
|---|
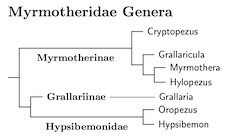
Antpitta Subfamilies: I've used three subfamilies to organize the antpittas: Myrmotherinae, Grallariinae, and Hypsibemonidae. Each corresponds to a split that occurred over 20 million years ago. I have not added tribes, which could be reasonably assigned to coincide with the genera.
Genus changes: I have made several changes to the genera in order to make sure that the deepest divisions are reflected in the generic structure as much as possible. More precisely, each side of any division deeper than 16 mya gets at least one genus. In fact, there is only one genus that spans a division deeper than 13 mya (Hylopezus).
To accomplish this, we have transferred three Hylopezus antpittas to Myrmothera, many of the Grallaria are now in Hypsibemon (Cabanis 1847, type ruficapilla), and the first four Hypsibemon have been moved to Oropezus.
Speckle-breasted Antpitta: Based on Carneiro, Bravo and Aleixo (2019), the Speckle-breasted Antpitta, Hylopezus nattereri, has been transferred to the genus Cryptopezus. It is the basal branch in Myrmotherinae. It's common ancestor with the rest of Myrmotherinae lived roughly 19.5 mya according to Harvey et al. (2020).
Rusty-breasted Antpitta: Based on Van Doren et al. (2018) and SACC #784 and SACC #801, the Rusty-breasted Antpitta, Grallaricula ferrugineipectus, is split into:
- Rusty-breasted Antpitta, Grallaricula ferrugineipectus
- Rufous-breasted Antpitta, Grallaricula leymebambae
Thrush-like Antpitta: Based on Carneiro et al. (2018) and SACC #785, the Thrush-like Antpitta, Myrmothera campanisona, has been split into:
- Thrush-like Antpitta, Myrmothera campanisona
- Tapajos Antpitta, Myrmothera subcanescens
Spotted Antpitta: It has been suspected that the Spotted Antpitta is more than one species (Ridgely and Tudor, 1994; Hilty, 2003; Krabbe and Schulenberg, 2003). Carneiro et al. (2012) made a close study of this. Based on their results, the Spotted Antpitta, Hylopezus macularius, has been split into three species:
- Spotted Antpitta, Hylopezus macularius, including dilutus
- Alta Floresta Antpitta, Hylopezus whittakeri
- Snethlage's Antpitta, Hylopezus paraensis
The subspecies names differ a bit from HBW-8 (Krabbe and Schulenberg, 2003), but match the HBW Checklist.
Carneiro et al. (2012) suggested one more split, Zimmer's Antpitta, Hylopezus dilutus. They resurrected the name dilutus, previously considered a junior synonym of paraensis. They also submerged diversus into dilutus. However, it remains unsupported by sufficient genetic or vocal evidence, and we will pass on that one pending additional information. See also see SACC #622, which also describes the ranges of the three species.
Variegated Antpitta: Based on Harvey et al. (2020) and the HBW Checklist, the Variegated Antpitta, Grallaria varia, has been split into:
- Northern Variegated Antpitta, Grallaria varia, including cinereiceps and distincta
- Southern Variegated Antpitta, Grallaria imperator, including intercedens
Rufous Antpitta complex: The Rufous Antpitta project has finally paid off. Based on SACC #883, Chesser et al. (2020a) and Isler et al. (2020) the Rufous Antpitta complex (Rufous, Bicolored, Chestnut) is split as follows: Chestnut Antpitta, Oropezus blakei, is split into:
- Oxapampa Antpitta, Oropezus centralis
- Ayacucho Antpitta, Oropezus ayacuchensis
- Chestnut Antpitta, Oropezus blakei
Rufous Antpitta, Oropezus rufulus, is split into 12 species:
- Sierra Nevada Antpitta, Oropezus spatiator
- Perija Antpitta, Oropezus saltuensis
- Muisca Antpitta, Oropezus rufulus
- Urubamba Antpitta, Oropezus occabambae, including marcapatensis
- Puno Antpitta, Oropezus sinaensis
- Bolivian Antpitta, Oropezus cochabambae
- Cajamarca Antpitta, Oropezus cajamarcae
- Chami Antpitta, Oropezus alvarezi
- Equatorial Antpitta, Oropezus saturatus
- Graves's Antpitta, Oropezus gravesi
- O'Neill's Antpitta, Oropezus oneilli
- Junin Antpitta, Oropezus obscurus
This involved the naming of several new taxa: alvarezi (Cuervo, Cadena, Isler, & Chesser), gravesi (Isler, Chesser, Robbins & Hosne), oneilli (Chesser & Isler), centralis and ayacuchensis (Hosner, Robbins, Isler, & Chesser), marcapatensis (Isler & Chesser), and sinaensis (Robbins, Isler, Chesser, & Tobias).
Urrao Antpitta: The Urrao Antpitta, Grallaria urraoensis, has been named twice. The first formal description was by Barrera and Bartels (2010) using the name Grallaria fenwickorum. The SACC considers this description inadequate, (SACC #479B). They rely on the description by the original discoverers in Carantón and Certuche (2010), who use the name Grallaria urraoensis. There are allegations and controversy surrounding the unfortunate circumstances that led to the two different names. I won't further comment on that. Normally, the name Grallaria fenwickorum would have priority, being published first. However, since the SACC considers the proposed type speciment to be inadequate to identify the bird, they use the second published name, urraoensis. I follow the SACC decision in this matter.
Myrmotheridae: Antpittas MacGillivray, 1839
7 genera, 69 species Not HBW Family
Myrmotherinae MacGillivray, 1839
- Speckle-breasted Antpitta, Cryptopezus nattereri
- Sucre Antpitta, Grallaricula cumanensis
- Slate-crowned Antpitta / Slaty-crowned Antpitta, Grallaricula nana
- Rusty-breasted Antpitta, Grallaricula ferrugineipectus
- Crescent-faced Antpitta, Grallaricula lineifrons
- Rufous-breasted Antpitta, Grallaricula leymebambae
- Ochre-breasted Antpitta, Grallaricula flavirostris
- Peruvian Antpitta, Grallaricula peruviana
- Hooded Antpitta, Grallaricula cucullata
- Scallop-breasted Antpitta, Grallaricula loricata
- Ochre-fronted Antpitta, Grallaricula ochraceifrons
- White-lored Antpitta, Myrmothera fulviventris
- Amazonian Antpitta, Myrmothera berlepschi
- Thicket Antpitta, Myrmothera dives
- Thrush-like Antpitta, Myrmothera campanisona
- Tepui Antpitta, Myrmothera simplex
- Tapajos Antpitta, Myrmothera subcanescens
- Masked Antpitta, Hylopezus auricularis
- White-browed Antpitta, Hylopezus ochroleucus
- Streak-chested Antpitta, Hylopezus perspicillatus
- Spotted Antpitta, Hylopezus macularius
- Alta Floresta Antpitta, Hylopezus whittakeri
- Snethlage's Antpitta, Hylopezus paraensis
Grallariinae P.L. Sclater, 1890
- Undulated Antpitta, Grallaria squamigera
- Great Antpitta, Grallaria excelsa
- Giant Antpitta, Grallaria gigantea
- Gray-naped Antpitta, Grallaria griseonucha
- Plain-backed Antpitta, Grallaria haplonota
- Moustached Antpitta, Grallaria alleni
- Scaled Antpitta, Grallaria guatimalensis
- Tachira Antpitta, Grallaria chthonia
- Northern Variegated Antpitta, Grallaria varia
- Southern Variegated Antpitta, Grallaria imperator
Hypsibemonidae Sundevall, 1872
- Stripe-headed Antpitta, Oropezus andicolus
- Sierra Nevada Antpitta, Oropezus spatiator
- Perija Antpitta, Oropezus saltuensis
- Bicolored Antpitta, Oropezus rufocinereus
- Muisca Antpitta, Oropezus rufulus
- Oxapampa Antpitta, Oropezus centralis
- Ayacucho Antpitta, Oropezus ayacuchensis
- Urubamba Antpitta, Oropezus occabambae
- Puno Antpitta, Oropezus sinaensis
- Bolivian Antpitta, Oropezus cochabambae
- Cajamarca Antpitta, Oropezus cajamarcae
- Chami Antpitta, Oropezus alvarezi
- Equatorial Antpitta, Oropezus saturatus
- Chestnut Antpitta, Oropezus blakei
- Graves's Antpitta, Oropezus gravesi
- O'Neill's Antpitta, Oropezus oneilli
- Junin Antpitta, Oropezus obscurus
- Elusive Antpitta, Hypsibemon eludens
- Ochre-striped Antpitta, Hypsibemon dignissimus
- Jocotoco Antpitta, Hypsibemon ridgelyi
- Chestnut-naped Antpitta, Hypsibemon nuchalis
- Pale-billed Antpitta, Hypsibemon carrikeri
- White-throated Antpitta, Hypsibemon albigula
- Chestnut-crowned Antpitta, Hypsibemon ruficapilla
- Watkins's Antpitta, Hypsibemon watkinsi
- Yellow-breasted Antpitta, Hypsibemon flavotinctus
- Santa Marta Antpitta, Hypsibemon bangsi
- Cundinamarca Antpitta, Hypsibemon kaestneri
- Urrao Antpitta, Hypsibemon urraoensis
- Brown-banded Antpitta, Hypsibemon milleri
- Tawny Antpitta, Hypsibemon quitensis
- White-bellied Antpitta, Hypsibemon hypoleucus
- Red-and-white Antpitta, Hypsibemon erythroleucus
- Rufous-faced Antpitta, Hypsibemon erythrotis
- Rusty-tinged Antpitta, Hypsibemon przewalskii
- Bay Antpitta, Hypsibemon capitalis
Rhinocryptidae: Tapaculos Wetmore, 1926 (1837)
Notes — Rhinocryptidae
 |
| Click for Rhinocryptidae species |
|---|
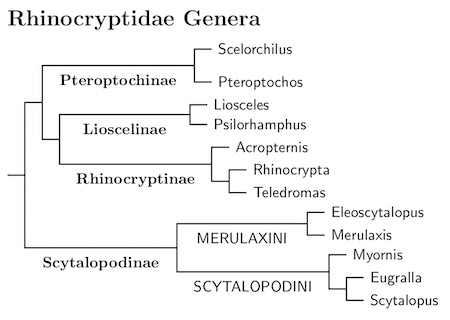
Tapaculo Subfamilies: The overall structure of the Rhinocryptidae comes from Harvey et al. (2020). There are several deep division within Rhinocryptidae. The subfamilies Pteroptochinae, Lioscelinae, Rhinocryptinae, and Scytalopodinae represent the deepest divisions in the tapaculo family. According to Harvey et al., these occurred between 20 and 24 million years ago.
I also divided the relatively speciose subfamily Scytalopodinae into two tribes, Merulaxini and Scytalopodini, each with an age of about 17.4 million years. I did not divide the smaller subfamiles into tribes. There's no deep division within Pteroptochinae. Tribes coincide with genera in the other two subfamilies.
When considering species, the Harvey et al. (2020) phylogeny has been augmented by Cadena et al. (2020), Krabbe et al. (2020), and Stiles et al. (2017) to include all of the currently recognized Scytalopus.
The relatively narrow age range for the subfamilies has made it a little difficult to place them correctly. The same groups appeared in Maurício et al. (2008), Moyle et al. (2009), and Ericson et al. (2010), but in somewhat different order. The paper by Ericson et al. was important as it put together all of the genera that comprise the Rhinocryptidae, excluding all pretenders such as the crescent-chests.
An alternative is the morphological phylogeny used by Maurício et al. (2012) which does not match the Harvey et al. phylogeny used here (nor that of Moyle et al., 2009; nor Ericson et al., 2010). I consider that an artifact of their morphological analysis, and find it unsurprising. One big problem with using morphology is that the characters used in the analysis are subject to a lot of convergence for functional reasons even if they are genetically different.
Eleoscytalopus: In version 2.04, the White-breasted and Bahia Tapaculos were moved out of Scytalopus into a new genus, Eleoscytalopus (see Maurício et al., 2008).
Scytalopus: The removal of Eleoscytalopus makes Scytalopus a recent radiation. The common ancestor of the Scytalopus tapaculos lived a mean 6.5 mya. In that 6.5 million years, the ancestral Scytalopus has turned into at least 49 species, and there are more to be added once the fieldwork is done. This genus is one of the great speciators!
Boa Nova Tapaculo: Based on Maurício et al. (2014), the Boa Nova Tapaculo, Scytalopus gonzagai, has been split from the Mouse-colored Tapaculo, Scytalopus speluncae.
Diamantina Tapaculo: The Diamantina Tapaculo, Scytalopus diamantinensis, was added to the list in 2.05. The composition of Scytalopus is currently very contentious. The SACC discussion of the Diamantina Tapaculo demonstrates how complex the issues are.
Rock Tapaculo: The Rock Tapaculo, Scytalopus petrophilus, was noted by Bornschein et al. (2007) and included in the genetic analysis of Mata et al. (2009). It was formally described by Whitney et al. (2010).
Names in the speluncae complex: The names of these taxa are in dispute, with some (Raposo et al., 2006, 2008) contending that speluncae does not apply to the Mouse-colored Tapaculo, but that it may apply to petrophilus. Raposo et al. (2006) introduced the alternate name notorius for the Mouse-colored Tapaculo. Maurício et al. (2010) argue in favor of the standard treatment, which is followed by the SACC and here.
To clarify a bit, regardless of the scientific names, the Mouse-colored Tapaculo (aka notorius) is the dark gray species (or species group) of more coastal mountains from Rio Grande do Sul to at least Minas Gerais and Espírito Santo, including the Serra do Caparaó, Serra do Mar, and Serra da Mantiqueira, while the Rock Tapaculo (aka petrophilus) is the light gray form in the interior range, Serra do Espinhaço.
The issue concerning the name is that the type specimen of speluncae was supposedly collected at São João del Rei, which is Rock Tapaculo territory. However, there are doubts whether this is correct, and others argue that in fact the specimen is a Mouse-colored Tapaculo with the actual collection location unknown. The specimen, collected by Ménétriés, is not in good shape, and access to it has been limited. The issues are whether Ménétriés was in error about the location for this specimen, and whether it is in fact a Mouse-colored Tapaculo. I'm following the SACC here, not because I'm entirely convinced they've made the right decision, but because to do otherwise would only futher complicate the issue.
New Scytalopus Tapaculo Species: All three are described in Krabbe et al. (2020), which also provides a phylogenetic analysis. See also SACC #852, SACC #853, and SACC #854, respectively.
- White-winged Tapaculo, Scytalopus krabbei
- Jalca Tapaculo, Scytalopus frankeae
- Ampay Tapaculo, Scytalopus whitneyi
Paramo Tapaculo: Based on Krabbe and Cadena (2010), Cadena et al. (2020), Krabbe et al. (2020), and SACC #855, the Paramo Tapaculo, Scytalopus opacus, is split into:
- Paramo Tapaculo, Scytalopus opacus
- Loja Tapaculo, Scytalopus androstictus
See also SACC #446.
Tatama Tapaculo / Alto Pisones Tapaculo: The “Alto Pisones” Tapaculo was finally described by Stiles et al. (2017). I have added it to the TiF list using their suggested English name: Tatama Tapaculo, Scytalopus alvarezlopezi, but have kept Alto Pisones as an alternate name. It is sister to the Ecuadorian Tapaculo / El Oro Tapaculo, Scytalopus robbinsi.
Perija Tapaculo: The Perija Tapaculo, Scytalopus perijanus, was described in Avendaño et al., (s2015).
Blackish Tapaculo: Based on Cadena et al. (2020) and and SACC #858, the Blackish Tapaculo, Scytalopus latrans, is split into:
- Blackish Tapaculo, Scytalopus latrans
- Utcubamba Tapaculo, Scytalopus intermedius
Junin Tapaculo: The newly described Junin Tapaculo, Scytalopus gettyae, has been added (see Hosner et al., 2013). This bird has been known of for a while as the Satipo form of Large-footed Tapaculo and is not to be confused with the so-called Millpo Tapaculo, which occurs higher up. It appears to be part of the latrans group.
Rhinocryptidae: Tapaculos Wetmore, 1926 (1837)
12 genera, 65 species HBW-8
Pteroptochinae P. L. Sclater, 1858 (1852)
- White-throated Tapaculo, Scelorchilus albicollis
- Chucao Tapaculo, Scelorchilus rubecula
- Moustached Turca, Pteroptochos megapodius
- Black-throated Huet-huet, Pteroptochos tarnii
- Chestnut-throated Huet-huet, Pteroptochos castaneus
Lioscelinae Mauricio et al. 2012
- Rusty-belted Tapaculo, Liosceles thoracicus
- Spotted Bamboowren, Psilorhamphus guttatus
Rhinocryptinae Wetmore, 1926 (1837)
- Ocellated Tapaculo, Acropternis orthonyx
- Crested Gallito, Rhinocrypta lanceolata
- Sandy Gallito, Teledromas fuscus
Scytalopodinae J. Müller, 1846
Merulaxini Mauricio et al., 2012
- White-breasted Tapaculo, Eleoscytalopus indigoticus
- Bahia Tapaculo, Eleoscytalopus psychopompus
- Slaty Bristlefront, Merulaxis ater
- Stresemann's Bristlefront, Merulaxis stresemanni
Scytalopodini J. Müller, 1846
- Ash-colored Tapaculo, Myornis senilis
- Ochre-flanked Tapaculo, Eugralla paradoxa
- Marsh Tapaculo, Scytalopus iraiensis
- Mouse-colored Tapaculo, Scytalopus speluncae
- Boa Nova Tapaculo, Scytalopus gonzagai
- Brasilia Tapaculo, Scytalopus novacapitalis
- Diamantina Tapaculo, Scytalopus diamantinensis
- Planalto Tapaculo, Scytalopus pachecoi
- Rock Tapaculo, Scytalopus petrophilus
- Dusky Tapaculo, Scytalopus fuscus
- Magellanic Tapaculo, Scytalopus magellanicus
- Ancash Tapaculo, Scytalopus affinis
- White-winged Tapaculo, Scytalopus krabbei
- Paramillo Tapaculo, Scytalopus canus
- Paramo Tapaculo, Scytalopus opacus
- Loja Tapaculo, Scytalopus androstictus
- White-browed Tapaculo, Scytalopus superciliaris
- Zimmer's Tapaculo, Scytalopus zimmeri
- Puna Tapaculo, Scytalopus simonsi
- Neblina Tapaculo, Scytalopus altirostris
- Jalca Tapaculo, Scytalopus frankeae
- Ampay Tapaculo, Scytalopus whitneyi
- Vilcabamba Tapaculo, Scytalopus urubambae
- Diademed Tapaculo, Scytalopus schulenbergi
- Trilling Tapaculo, Scytalopus parvirostris
- Bolivian Tapaculo / Bolivian White-crowned Tapaculo, Scytalopus bolivianus
- Santa Marta Tapaculo, Scytalopus sanctaemartae
- White-crowned Tapaculo / Northern White-crowned Tapaculo, Scytalopus atratus
- Narino Tapaculo, Scytalopus vicinior
- Silvery-fronted Tapaculo, Scytalopus argentifrons
- Tacarcuna Tapaculo, Scytalopus panamensis
- Choco Tapaculo, Scytalopus chocoensis
- Magdalena Tapaculo, Scytalopus rodriguezi
- Stiles's Tapaculo, Scytalopus stilesi
- Tatama Tapaculo / Alto Pisones Tapaculo, Scytalopus alvarezlopezi
- Ecuadorian Tapaculo / El Oro Tapaculo, Scytalopus robbinsi
- Merida Tapaculo, Scytalopus meridanus
- Caracas Tapaculo, Scytalopus caracae
- Brown-rumped Tapaculo, Scytalopus latebricola
- Perija Tapaculo, Scytalopus perijanus
- Pale-bellied Tapaculo, Scytalopus griseicollis
- Spillmann's Tapaculo, Scytalopus spillmanni
- Chusquea Tapaculo, Scytalopus parkeri
- Long-tailed Tapaculo, Scytalopus micropterus
- Rufous-vented Tapaculo, Scytalopus femoralis
- Blackish Tapaculo, Scytalopus latrans
- Tschudi's Tapaculo, Scytalopus acutirostris
- Unicolored Tapaculo, Scytalopus unicolor
- Utcubamba Tapaculo, Scytalopus intermedius
- Large-footed Tapaculo, Scytalopus macropus
- Junin Tapaculo, Scytalopus gettyae
Formicariidae: Antthrushes G.R. Gray, 1840 (1825)
Notes — Formicariidae
 |
| Click for Formicariidae species |
|---|
The Antthrushes or Formicariidae are the last family before the Ovenbirds. We need to ask whether the Antthrushes belong with the Ovenbirds. The answer is no. The split between the Antthrushes and Ovenbirds dates to 32.5 mya, according to Harvey et al. (2020). All of the Ovenbirds share a common ancestor at about 21.3 mya. With such a gap, it would make no sense to combine them.
Antthrush Subfamilies: The structure of the Formicariidae is quite simple. There are two genera, Formicarius and Chamaeza. But these two genera are separated by 23.2 million years! It makes sense to rank them as subfamilies: Formicariinae and Chamaezinae. One thing that distinguishes them are the smooth colors on the underparts of the Formicariinae and the patterns (scallops, bars, etc.) under the Chamaezinae.
Black-faced Antthrush: Based on Harvey et al. (2020), the NACC decision 2020-B11 on the Mayan Antthrush, and the HBW Checklist, the Black-faced Antthrush, Formicarius analis is split into:
- Mayan Antthrush, Formicarius moniliger, including pallidus and intermedius
- Northern Black-faced Antthrush, Formicarius hoffmanni, including umbrosus, panamensis, virescens, and griseoventris
- Southern Black-faced Antthrush, Formicarius analis, including connectens, zamorae, crissalis, and paraensis
Striated Antthrush: Based on Harvey et al. (2020) and the HBW Checklist, the Striated Antthrush, Chamaeza nobilis is split into:
- Western Striated Antthrush, Chamaeza nobilis, including rubida
- Eastern Striated Antthrush, Chamaeza fulvipectus
Short-tailed Antthrush: Based on Harvey et al. (2020) and the HBW Checklist, which notes 5 song grous, the Short-tailed Antthrush, Chamaeza campanisona is split into:
- Brazilian Short-tailed Antthrush, Chamaeza campanisona, including tshororo
- Roraiman Short-tailed Antthrush, Chamaeza fulvescens, including obscura, yavii, and huachamacarii
- Colombian Short-tailed Antthrush, Chamaeza columbiana, including punctigula
- Venezuelan Short-tailed Antthrush, Chamaeza venezuelana
- Peruvian Short-tailed Antthrush, Chamaeza olivacea, including berlepschi and boliviana
Formicariidae: Antthrushes G.R. Gray, 1840 (1825)
2 genera, 18 species HBW-8
Formicariinae: Smooth Antthrushes G.R. Gray, 1840 (1825)
- Rufous-fronted Antthrush, Formicarius rufifrons
- Rufous-capped Antthrush, Formicarius colma
- Rufous-breasted Antthrush, Formicarius rufipectus
- Mayan Antthrush, Formicarius moniliger
- Black-headed Antthrush, Formicarius nigricapillus
- Northern Black-faced Antthrush, Formicarius hoffmanni
- Southern Black-faced Antthrush, Formicarius analis
Chamaezinae: Patterned Antthrushes Informal
- Rufous-tailed Antthrush, Chamaeza ruficauda
- Such's Antthrush / Cryptic Antthrush, Chamaeza meruloides
- Barred Antthrush, Chamaeza mollissima
- Schwartz's Antthrush, Chamaeza turdina
- Brazilian Short-tailed Antthrush, Chamaeza campanisona
- Western Striated Antthrush, Chamaeza nobilis
- Eastern Striated Antthrush, Chamaeza fulvipectus
- Roraiman Short-tailed Antthrush, Chamaeza fulvescens
- Colombian Short-tailed Antthrush, Chamaeza columbiana
- Venezuelan Short-tailed Antthrush, Chamaeza venezuelana
- Peruvian Short-tailed Antthrush, Chamaeza olivacea
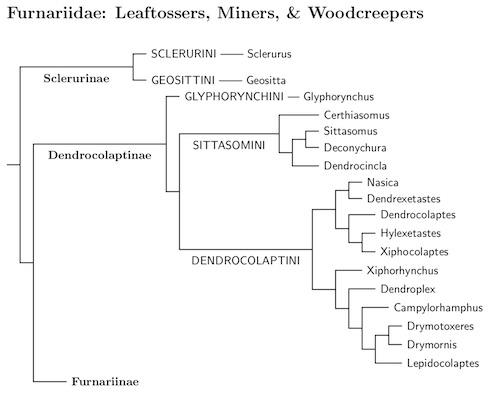
Furnariidae: Ovenbirds G.R. Gray, 1840
72 genera, 346 species HBW-8
 At the family level, the taxonomy agrees with the SACC, folding the
woodcreepers into the Furnariidae as the subfamily Dendrocolaptinae rather
than maintaining them as a separate family. That arrangement did not work
because the woodcreepers are nested within the Furnariidae, as shown on the
diagram to the right. The Furnariidae consists of three subfamilies:
Sclerurinae (leaftossers and miners), Furnariinae (ovenbirds), and
Dendrocolaptinae (woodcreepers). I arrange them as Sclerurinae,
Dendrocolaptinae, Furnariinae because Furnariinae is much larger than
Dendrocolaptinae.
At the family level, the taxonomy agrees with the SACC, folding the
woodcreepers into the Furnariidae as the subfamily Dendrocolaptinae rather
than maintaining them as a separate family. That arrangement did not work
because the woodcreepers are nested within the Furnariidae, as shown on the
diagram to the right. The Furnariidae consists of three subfamilies:
Sclerurinae (leaftossers and miners), Furnariinae (ovenbirds), and
Dendrocolaptinae (woodcreepers). I arrange them as Sclerurinae,
Dendrocolaptinae, Furnariinae because Furnariinae is much larger than
Dendrocolaptinae.
| Group | Age |
|---|---|
| Melanopareiidae | 35.0 |
| Conopophagidae | 33.8 |
| Thamnophilidae | 33.8 |
| Myrmotheridae | 35.2 |
| Rhinocryptidae | 33.5 |
| Formicariidae | 32.5 |
| Furnariidae (broad) | 32.5 |
| Sclerurinae | 21.3 |
| Dendrocolaptinae | 18.7 |
| Furnariinae | 18.7 |
If the woodcreepers were separated, the Furnariidae would no longer be monophyletic. The leaftossers and miners would also have to be separated. This could be done by promoting all three subfamilies to families. However, these families would be much younger than the other families in Furnariida. The table to the right gives the ages, in millions of years for the Furnariid families and the three groups in question.
Sibley and Monroe's attempt to base the level of each taxon on their age convinced me that a using fixed age scale for the entire avian tree is a very foolish approach. They demonstrate that it just doesn't do of good job of handling the variable diversity that we face when classifying birds. Think of their Fringillidae — a single family containing a ninth of all bird species. Nonetheless, I do think it reasonable to do so locally. In this case, locally means within the Furnariida, so we have one family Furnariidae, comprised of three subfamilies: Sclerurinae, Dendrocolaptinae, and Furnariinae.
 |
| Click for Sclerurinae-Dendrocolaptinae species tree |
|---|
Notes — Leaftossers and Miners
Sclerurinae: The two genera in Sclerurinae, leaftossers and miners, are quite different. Even a beginner will not mistake one for the other. They are also separated by about 17.7 million years of evolution, according to Harvey et al. (2020). To emphasize this, I have placed them in separate tribes, as has been done for other such taxa. The tribes are Sclerurinini (leaftossers) and Geosittini (miners).
Tawny-throated Leaftosser: Based on Harvey et al. (2020), the HBW Checklist, d'Horta et al. (2013), and Cooper and Cuervo (2017), the Tawny-throated Leaftosser, Sclerurus mexicanus, is split into:
- Mexicqn Leaftosser, Sclerurus mexicanus
- Dusky Leaftosser, Sclerurus pullus
- Pacific Leaftosser, Sclerurus obscurior
- Andean Leaftosser, Sclerurus andinus
- Amazonian Leaftosser, Sclerurus macconnelli, including peruvianus, macconnelli, and bahiae
The SACC currently considers this complex two species with the South Amerian birds collectively called South American Leaftosser. See the SACC Proposals 603, 752, and 860.
Sclerurinae: Leaftossers and Miners Swainson, 1827
Sclerurini: Leaftossers Swainson 1827
- Scaly-throated Leaftosser, Sclerurus guatemalensis
- Black-tailed Leaftosser, Sclerurus caudacutus
- Gray-throated Leaftosser, Sclerurus albigularis
- Rufous-breasted Leaftosser, Sclerurus scansor
- Mexican Leaftosser, Sclerurus mexicanus
- Dusky Leaftosser, Sclerurus pullus
- Short-billed Leaftosser, Sclerurus rufigularis
- Pacific Leaftosser / Dusky Leaftosser, Sclerurus obscurior
- Andean Leaftosser, Sclerurus andinus
- Amazonian Leaftosser, Sclerurus macconnelli
Geosittini: Miners Informal
- Coastal Miner, Geositta peruviana
- Slender-billed Miner, Geositta tenuirostris
- Common Miner, Geositta cunicularia
- Puna Miner, Geositta punensis
- Campo Miner, Geositta poeciloptera
- Thick-billed Miner, Geositta crassirostris
- Rufous-banded Miner, Geositta rufipennis
- Grayish Miner, Geositta maritima
- Dark-winged Miner, Geositta saxicolina
- Creamy-rumped Miner, Geositta isabellina
- Short-billed Miner, Geositta antarctica
Notes — Woodcreepers
Woodcreeper Tribes: We now turn to the woodcreepers. I've divided the woodcreeper subfamily Dendrocolaptinae into three tribes: Glyphorynchini (for the Wedge-billed Woodcreeper), Sittasomini, and Dendrocolaptini. The boundaries of these groups have been generally stable, although there is some disagreement abour their relationships. Of course, the current version of the TiF list follows Harvey et al. (2020).
This arrangment has the tribe Glyphorynchini as basal woodcreeper, increasing the case that it really is basal. This concurs with Derryberry et al. (2012), but disagrees with Derryberry et al. (2011), Moyle et al. (2009b), and Ohlsen et al. (2013), which have it sister to Sittasomini.
Irestedt et al. (2009b), Derryberry et al. (2011, 2012), and Moyle et al. (2009b) can be read as concurring that Sittasomini and Dendrocolaptini are natural groups. They agree on the membership and even the substructure of the Dendrocolaptini. They are all consistent with the same genera, something only accomplished in the last few years.
Both the Sittasomini and Dendrocolaptini and have been considered natural groups on morphological grounds too. There is an interesting discussion of this in Marantz et al. (2003, aka HBW-8), which distinguishes a group of “strong-billed” woodcreepers, meaning the Dendrocolaptini and “intermediate” woodcreepers (Glyphorynchini and Sittasomini), which have some traits of the other ovenbirds that are not shared by the strong-billed woodcreepers. In other words, they are morphologically intermediate between the other ovenbirds and the strong-billed woodcreepers.
The big disagreement concerns the position of Glyphorynchus. Irestedt et al. and Derryberry et al. (2012) have it in the basal position, while Derryberry et al. (2011), Moyle et al. (2009b), and Ohlsen et al. (2013) have it sister to Dendrocolaptini. What can one conclude? Since Harvey et al. (2020) also endorse the basal position, I've gone with that.
Within Sittasomini, Deconychura has been split into two (Derryberry et al., 2010a). As a result the Spot-throated Woodcreeper is now Certhiasomus stictolaemus. It is basal in Sittasomini.
Long-tailed Woodcreeper: Based on Harvey et al. (2020), Barbosa (2010), and the HBW Checklist, the Long-tailed Woodcreeper, Deconychura longicauda is split into:
- Little Long-tailed Woodcreeper, Deconychura typica, including darienensis and minor
- Northern Long-tailed Woodcreeper, Deconychura longicauda
- Southern Long-tailed Woodcreeper, Deconychura pallida, including connectens and zimmeri
This complex also contains an undescribed species from the east Andes of Peru (Barbosa, 2010). Not only was it sampled by Barbosa, but apparently also by Harvey et al. (2020).
Olivaceous Woodcreeper: Based on Harvey et al. (2020), the HBW Checklist, and Boseman (2006), the Olivaceous Woodcreeper, Sittasomus sylviellus is split into:
- Amazonian Woodcreeper, Sittasomus amazonus, including axillaris, transitivus, and viridis
- Olivaceous Woodcreeper, Sittasomus sylviellus, including olivaceus
- Reiser's Woodcreeper, Sittasomus reiseri
- Chaco Woodcreeper, Sittasomus griseicapillus
- Grayish Woodcreeper, Sittasomus griseus, including jaliscensis, sylvioides, gracileus, perijanus, and tachirensis
- Pacific Woodcreeper, Sittasomus aequatorialis
Plain-brown Woodcreeper complex: Based on Schultz et al. (2019) and Weir and Price (2011), with some help from Harvey et al. (2020) and the HBW Checklist, the Plain-brown Woodcreeper complex has been reorganized. The complex, which included only three species (Plain-winged, Tawny-winged, Plain-brown) in H&M-4, is now comprised of four species:
- Tawny-winged Woodcreeper, Dendrocincla anabatina, including typhla and saturata
- Plain-brown Woodcreeper, Dendrocincla fuliginosa, including rufoolivacea, trumaii (deprecated); ridgwayi, lafresnayei (both parts); meruloides, barinensis, deltana; phaeochroa, and neglecta (deprecated)
- d'Orbigny's Woodcreeper, Dendrocincla atrirostris
- Plain-winged Woodcreeper, Dendrocincla turdina, including taunayi
The subspecies taunayi is transferred to the Plain-winged Woodcreeper, Dendrocincla turdina.
Schultz et al. (2019) carried out extensive genetic testing of many individuals from this complex. They have carefully sorted out the subspecies of D. meruloides (see their Figure 2 and associated text). The subspecies lafresnayei is paraphyletic. That will have to be dealt with at some point. The subspecies can be grouped. The ridgwayi group also contains part of lafresnayei and the meruloides group also contains barinensis, deltana, and the remainder of lafresnayei. The third group is phaeochroa including neglecta. They could not identify a separate lineage for neglecta and it is perhaps best sunk into phaeochroa.
At this point we have 7 potential species-level taxa, the groups: atrirostris, turdina, fuliginosa, anabatina, meruloides, ridgwayi, and phaeochroa. So how do they fit together?
There's disagreement between the mitochondrial DNA tree and the multilocus tree. I follow the multilocus tree. In that case, lacking other data, I apply a 2 million year cutoff for species status. That yields 4 species, the groups anabatina, atrirostris, and turdina, and the combined group consisting of the fuliginosa, meruloides, ridgwayi, and phaeochroa groups. The name fuliginosa has priority. We also need an English name for atrirostris. Cory and Hellmayr (1925) called it d'Orbigny's Woodcreeper.
I have left the question of species status of ridgwayi, and phaeochroa for another day.
Finally, for those who need to revise their lists, or consider that there might be further splits, I draw your attention to range maps in Figures 5 and 6 in Schultz et al. (2019).
Red-billed Woodcreeper: Harvey et al. (2020) and Derryberry et al. (2012) provide support for Silva and Oren's (1995) treatment of Hylexetastes as four species. Accordingly, Red-billed Woodcreeper, Hylexetastes perrotii, is split into:
- Red-billed Woodcreeper, Hylexetastes perrotii
- Uniform Woodcreeper, Hylexetastes uniformis
- Brigida's Woodcreeper, Hylexetastes brigidai
Strong-billed Woodcreeper: Based on Harvey et al. (2020) and the HBW Checklist, the Strong-billed Woodcreeper, Xiphocolaptes promeropirhynchus, is split into:
- Amazonian Strong-billed Woodcreeper, Xiphocolaptes orenocensis, including berlepschi, paraensis, carajaensis, and obsoletus
- Central American Strong-billed Woodcreeper, Xiphocolaptes emigrans, including omiltemensis, sclateri, costaricensis, and panamensis
- Andean Strong-billed Woodcreeper, Xiphocolaptes promeropirhynchus, including sanctaemartae, rostratus, virgatus, procerus, macarenae, neblinae, tenebrosus, ignotus, crassirostris, compressirostris, phaeopygus, lineatocephalus, and solivagus
Ocellated Woodcreeper: Based on Harvey et al. (2020), Sousa-Neves, Aleixo, and Sequeira (2013), and the HBW Checklist, the Ocellated Woodcreeper, Xiphorhynchus ocellatus is split into:
- Ocellated Woodcreeper, Xiphorhynchus ocellatus, including perplexus
- Line-crowned Woodcreeper, Xiphorhynchus beauperthuysii, including lineatocapilla
This builds on the previous split of Tschudi's Woodcreeper, Xiphorhynchus chunchotambo from Ocellated Woodcreeper. I found the map in Sousa-Neves et al. to be very helpful for understanding the distribution of this complex. Their area A is the Chestnut-rumped Woodcreeper, areas B and C, which are south of the Amazon are Ocellated Woodcreeper, area D is Line-crowned Woodcreeper (north of the Amazon and west of the Rio Negro), and area E and the unlabelled yellow zone is Tschudi's Woodcreeper. Both Line-crowned and Tschudi's are present in the cross-hatched area.
Buff-throated Woodcreeper complex: The situation with the Cocoa (Xiphorhynchus susurrans), Buff-throated (X. guttatus), and Lafresnaye's (X. guttatoides) Woodcreepers is now clearer due to Rocha et al. (2015) and Aleixo (2001). I currently recognize 3 species in the complex.
- Lafresnaye's Woodcreeper, Xiphorhynchus guttatoides, including dorgibnyanus, eytoni, gracilirostris, and vicinalis (south and west of the Amazon/Negro).
- Cocoa Woodcreeper, Xiphorhynchus susurrans (Central America, Colombian Andes, northern Venezuela).
- Buff-throated Woodcreeper, Xiphorhynchus guttatus, including connectens and polystictus (north and east of the Amazon/Negro, Atlantic Forest).
Harvey et al. (2020) and Rocha et al. (2015) have different takes on the relationships here. I've followed Rocha et al. as it includes all of the taxa in the complex. In that case, the Cocoa and Buff-throated Woodcreepers are sister species, with the Lafresnaye's complex more distantly related. Harvey et al. insert the Black-striped Woodcreeper into the mix, as sister to the Cocoa Woodcreeper. They did not include Lafresnaye's in their analysis. I've a question mark on the Black-striped in the species tree to indicate that there is a problem here.
The Lafresnaye's complex may contain two additional species. However, as Rocha et al. (2015) point out, it would be prudent to sample upriver before accepting further splits. The dark-billed subspecies east of the Xingu (eytoni and gracilirostris) are sister taxa and only distantly related to the other subspecies of Lafresnaye's. They have sometimes been united as Dusky-billed Woodcreeper with the other dark-billed subspecies, vicinalis. However, the sampled vicinalis are quite a bit more closely related to guttatoides than to eytoni or gracilirostris. The subspecies vicinalis actually ranges from the Madeira to the Xingu (not to the Tapajós, as previously thought). It has been thought that some are intergrades between eytoni and guttatoides, but that was not supported by Rocha et al.'s samples.
Curve-billed Scythebill: Based on Harvey et al. (2020), Aleixo et al. (2013), Portes et al. (2013), and the HBW Checklist, the Curve-billed Scythebill, Campylorhamphus procurvoides, is split into:
- Xingu Scythebill, Campylorhamphus multostriatus
- Tapajos Scythebill, Campylorhamphus probatus, including cardosoi
- Curve-billed Scythebill, Campylorhamphus procurvoides, including sanus and gyldenstolpei
Lineated Woodcreeper complex: Based on Rodrigues et al. (2013) and SACC proposals #620, #660, #868, and #894, the Lineated Woodcreeper, Lepidocolaptes albolineatus, has been is split into 4 species. They are:
- Guianan Woodcreeper, Lepidocolaptes albolineatus
- Duida Woodcreeper, Lepidocolaptes duidae
- Inambari Woodcreeper, Lepidocolaptes fatimalimae
- Dusky-capped Woodcreeper, Lepidocolaptes fuscicapillus, including layardi and madeirae
Dendrocolaptinae: Woodcreepers G.R. Gray, 1840
Glyphorynchini: Wedge-billed Woodcreeper Sharpe 1901
- Wedge-billed Woodcreeper, Glyphorynchus spirurus
Sittasomini: Intermediate Woodcreepers Ridgway 1911
- Spot-throated Woodcreeper, Certhiasomus stictolaemus
- Northern Long-tailed Woodcreeper, Deconychura longicauda
- Little Long-tailed Woodcreeper, Deconychura typica
- Southern Long-tailed Woodcreeper, Deconychura pallida
- Amazonian Woodcreeper, Sittasomus amazonus
- Olivaceous Woodcreeper, Sittasomus sylviellus
- Reiser's Woodcreeper, Sittasomus reiseri
- Chaco Woodcreeper, Sittasomus griseicapillus
- Grayish Woodcreeper, Sittasomus griseus
- Pacific Woodcreeper, Sittasomus aequatorialis
- Tyrannine Woodcreeper, Dendrocincla tyrannina
- White-chinned Woodcreeper, Dendrocincla merula
- Ruddy Woodcreeper, Dendrocincla homochroa
- Tawny-winged Woodcreeper, Dendrocincla anabatina
- Plain-brown Woodcreeper, Dendrocincla fuliginosa
- d'Orbigny's Woodcreeper, Dendrocincla atrirostris
- Plain-winged Woodcreeper, Dendrocincla turdina
Dendrocolaptini: Strong-billed Woodcreepers GR Gray 1840
- Long-billed Woodcreeper, Nasica longirostris
- Cinnamon-throated Woodcreeper, Dendrexetastes rufigula
- Northern Barred-Woodcreeper, Dendrocolaptes sanctithomae
- Amazonian Barred-Woodcreeper, Dendrocolaptes certhia
- Planalto Woodcreeper, Dendrocolaptes platyrostris
- Black-banded Woodcreeper, Dendrocolaptes picumnus
- Hoffmanns's Woodcreeper, Dendrocolaptes hoffmannsi
- Bar-bellied Woodcreeper, Hylexetastes stresemanni
- Red-billed Woodcreeper, Hylexetastes perrotii
- Uniform Woodcreeper, Hylexetastes uniformis
- Brigida's Woodcreeper, Hylexetastes brigidai
- White-throated Woodcreeper, Xiphocolaptes albicollis
- Amazonian Strong-billed Woodcreeper, Xiphocolaptes orenocensis
- Moustached Woodcreeper, Xiphocolaptes falcirostris
- Central American Strong-billed Woodcreeper, Xiphocolaptes emigrans
- Andean Strong-billed Woodcreeper, Xiphocolaptes promeropirhynchus
- Great Rufous Woodcreeper, Xiphocolaptes major
- Elegant Woodcreeper, Xiphorhynchus elegans
- Spix's Woodcreeper, Xiphorhynchus spixii
- Ceara Woodcreeper, Xiphorhynchus atlanticus
- Lesser Woodcreeper, Xiphorhynchus fuscus
- Chestnut-rumped Woodcreeper, Xiphorhynchus pardalotus
- Ocellated Woodcreeper, Xiphorhynchus ocellatus
- Tschudi's Woodcreeper, Xiphorhynchus chunchotambo
- Line-crowned Woodcreeper, Xiphorhynchus beauperthuysii
- Spotted Woodcreeper, Xiphorhynchus erythropygius
- Olive-backed Woodcreeper, Xiphorhynchus triangularis
- Striped Woodcreeper, Xiphorhynchus obsoletus
- Ivory-billed Woodcreeper, Xiphorhynchus flavigaster
- Black-striped Woodcreeper, Xiphorhynchus lachrymosus
- Lafresnaye's Woodcreeper, Xiphorhynchus guttatoides
- Cocoa Woodcreeper, Xiphorhynchus susurrans
- Buff-throated Woodcreeper, Xiphorhynchus guttatus
- Straight-billed Woodcreeper, Dendroplex picus
- Zimmer's Woodcreeper, Dendroplex kienerii
- Black-billed Scythebill, Campylorhamphus falcularius
- Brown-billed Scythebill, Campylorhamphus pusillus
- Xingu Scythebill, Campylorhamphus multostriatus
- Curve-billed Scythebill, Campylorhamphus procurvoides
- Red-billed Scythebill, Campylorhamphus trochilirostris
- Tapajos Scythebill, Campylorhamphus probatus
- Greater Scythebill, Drymotoxeres pucheranii
- Scimitar-billed Woodcreeper, Drymornis bridgesii
- Scalloped Woodcreeper, Lepidocolaptes falcinellus
- Scaled Woodcreeper, Lepidocolaptes squamatus
- Narrow-billed Woodcreeper, Lepidocolaptes angustirostris
- Duida Woodcreeper, Lepidocolaptes duidae
- Inambari Woodcreeper, Lepidocolaptes fatimalimae
- Dusky-capped Woodcreeper, Lepidocolaptes fuscicapillus
- Guianan Woodcreeper, Lepidocolaptes albolineatus
- Streak-headed Woodcreeper, Lepidocolaptes souleyetii
- Montane Woodcreeper, Lepidocolaptes lacrymiger
- White-striped Woodcreeper, Lepidocolaptes leucogaster
- Spot-crowned Woodcreeper, Lepidocolaptes affinis
Furnariinae: True Ovenbirds G.R. Gray, 1840
The Xenops Problem
I have redone the ovenbirds multiple times during TiF's history, and have always had a problem with the genus Xenops, usually ranking it as a subfamily: Xenopinae. The problem with Xenops has been a lack of clarity whether Xenops is closer to the woodcreepers or true ovenbirds.
Some gene trees put Xenops is sister to Dendrocolaptinae, while others put it sister to Furnariinae (see Fjeldså et al., 2005, 2007; Irestedt et al., 2009b; Moyle et al., 2009b; Derryberry et al., 2011). Fjeldså et al. (2005) tell a nice story that makes Xenops somewhat analogous to a piculet. I have some sympathy for this argument as they sometimes remind me of piculets. Moreover, the nuclear RAG-1 and RAG-2 genes that Moyle et al. rely on may change too slowly to resolve all of the find details of the tree. On the other hand, the other genes used by Derryberry et al. did not change the RAG picture. Further, the cytochrome-b tree in Fjeldså et al. (2005) puts Xenops with the Furnariinae. However, Fjeldså et al. (2005, 2007) both use additional genes and end up with Xenops near the woodcreepers. Irestedt et al. (2009b) agree in their overall tree, but the is considerable disagreement among the individual gene trees.
The newer analyses, including Derryberry et al. (2011) and Harvey et al. (2020) mostly place Xenops sister to the rest of the ovenbirds. This is not universal. Fjeldså et al. (2020) is an exception. My current take is that is a transitional group between the woodcreepers and the rest of the ovenbirds, and as such, naturallly groups with the ovenbirds. I think the balance of the genetic information supports this, and its time to declare this settled, or at least settled enough to rerank Xenopinae as the basal tribe in the ovenbird subfamily Furnariinae.
Plain Xenops: Based on Harvey and Brumfield (2015) and the HBW Checklist, the Plain Xenops, Xenops minutus, is split into:
- White-throated Xenops, Xenops minutus
- Northwestern Plain Xenops, Xenops mexicanus, includes ridgwayi, littoralis, olivaceus, and neglectus
- Southeastern Plain Xenops, Xenops genibarbis, includes remoratus, ruficaudus, obsoletus, and alagoanus
Rufous-tailed Xenops: There's one other complication with Xenops. The Rufous-tailed Xenops does not belong in Xenops (Moyle et al., 2009b; Irestedt et al., 2009b). Fortunately, there is an available genus name: Microxenops (Chapman 1914). Accordingly, the Rufous-tailed Xenops becomes Microxenops milleri and moves to Pygarrhichadini. The genus Megaxenops was also formerly considered part of this group, but Remsen (2003) had already noted that it is not closely related to Xenops. It's in Philydorini.
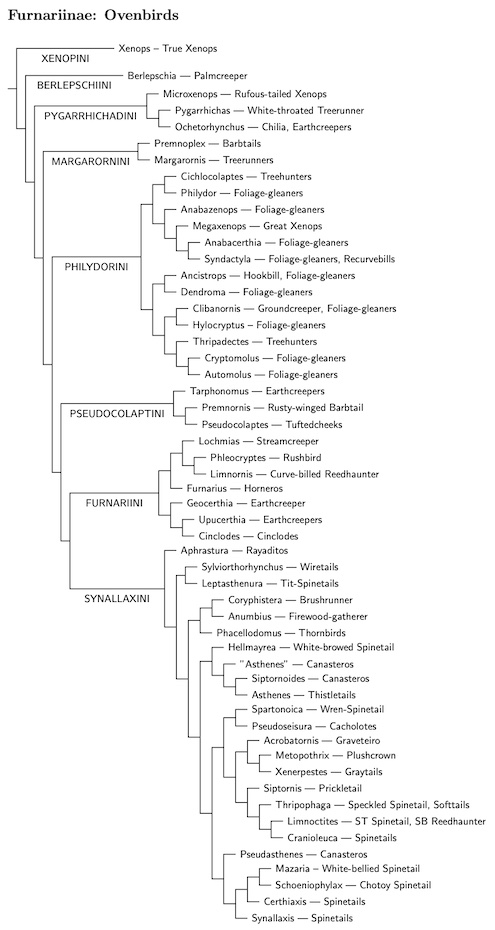
Ovenbird Tribes
I've changed the tribes a bit based on Harvey et al. (2020). As mentioned above, rather than treat Xenops as a subfamily, I have demoted it to a tribe in Furnariinae.
I've also split the tribe Furnariini into two tribes: Pseudocolaptini and Furnariini. Harvey et al. (2020) found that they form separate branches of the ovenbird tree, being part of a grade rather than being sister taxa. The Harvey et al. tree also does the same to Philydorini and Furnariini, and moves Margarornini to a more basal position.
The list here follows the recent comprehensive analysis of the suboscines by Harvey et al. (2020). As far as the ovenbirds are concerned, the differences between it and Derryberry et al. (2011) are small. Derryberry and 8 co-authors sampled all but 13 of the Furnariid species. Harvey et al. have done even better, with coverage of a number of possible splits and even some undescribed taxa.
The Harvey et al. topology should be seen as the culmination of a line of work that includes previous papers by Chesser et al. (2007), Fjeldså et al. (2005, 2007), Gonzalez and Wink (2008), Irestedt et al. (2004a, 2006a, 2009b), Moyle et al. (2009b), and Claramunt et al. (2010). Earlier versions of TiF used an overall arrangement is driven by the then recent papers by Irestedt et al. (2009b) and Moyle et al. (2009b).
I'm happy to say that, except for the changes mentioned above, I'm able to retain the same tribes (with the same members) as before Derryberry et al. The fact that this structure remains even though a number of new genes have been added to the analysis suggests that it is correct. There remain some minor issues concerning the arrangement of these tribes, both externally and internally, but the overall picture seems stable at this point.
 |
| Furnariinae genus tree based on Harvey et al. (2020). Click the diagram for the previous version based on Moyle et al. (2009b) and Derryberry et al. (2011). |
|---|
Following Xenopini, we have Berlepschia by itself as tribe Berlepschiini. It is followed by two more small tribe, Pygarrhichadini and Margarornini. Next comes the Furnariini-Philydorini group. According to Harvey et al. (2020), it's a grade of three tribes, but some earlier analyses suggested they may be sisters. Finally, we have the largest tribe of all, Synallaxini.
There have also been some changes within these groups, sometimes involving reoganized genera. Note that except for splits and lumps, the species composition of each tribe has remained virtually unchanged since version 2.13 of this page (June 30, 2009). However, some have been moved to different genera.
Notes — Philydorini
There are several changes within Philydorini resulting from Derryberry et al. (2011). One big one is the dismemberment of Philydor itself! It left only three species in Philydor. The other six were reallocated as follows:
- The Ochre-breasted and Rufous-tailed Foliage-gleaners moved to Anabacerthia
- I initially moved the Buff-fronted and Chestnut-winged Foliage-gleaners to Ancistrops. However, the differences between them and the Chestnut-winged Hookbill are rather large, so I've placed the two foliage-gleaners in the genus Dendroma (Swainson 1837, type rufa). See SACC #819. These are not that close to Philydor)
- The Rufous-rumped and Slaty-winged Foliage-gleaners first moved to Megaxenops. They didn't seem to fit comfortably with the Great Xenops and are now in Anabazenops.
Further, the two recurvebills Simoxenops have been moved into Syndactyla. They sit fairly deeply in Syndactyla, and even though they are distinctive, I don't see any other reasonable way to handle this.
The remaining changes to the Philydorini are confined to the Automolus branch. Chestnut-capped (Henna-capped) Foliage-gleaner is transferred from Hylocryptus to Clibanornis. The woodhaunteer genus Hyloctistes has been merged into Automolus. The Santa Marta Foliage-gleaner and the Ruddy Foliage-gleaner complex have been removed from Automolus and are now placed in Hylocryptus.
Cryptic Treehunter: The newly discovered (and possibly extinct) Cryptic Treehunter, Cichlocolaptes mazarbarnetti, has been added. See Mazar Barnett and Buzzetti (2014) and Claramunt (2014b).
Sharp-billed Treehunter: This has been moved from Heliobletus to Philydor. Also, note that one of the subspecies currently doesn't have a name! The problem is that contaminatus and camargoi refer to the same subspecies! See Penhallurick (2011) for details. For years we've heard there will be a replacement… we're still waiting.
Also, there seems to be a sister species in Bahia. It hasn't been described yet, but is included in Harvey et al. (2020).
Ruddy Foliage-gleaner complex: Both the Santa Marta Foliage-gleaner, Clibanornis rufipectus, and Henna-hooded Foliage-gleaner, Clibanornis erythrocephalus, are embedded in the complex. We First transfer all of these species to Hylocryptus (Chapman 1919, type erythrocephalus).
Based on Harvey et al. (2020), Claramunt et al. (2013), and the HBW Checklist, the Ruddy Foliage-gleaner, Hylocryptus rubiginosus, is split into:
- Rusty Foliage-gleaner, Hylocryptus rubiginosus, including guerrerensis, veraepacis, and fumosus
- Cinnamon-throated Foliage-gleaner, Hylocryptus cinnamomeigula
- Dusky Foliage-gleaner, Hylocryptus obscurus, including venezuelanus, caquetae, brunnescens,, and watkinsorum
- Black-tailed Foliage-gleaner, Hylocryptus nigricauda, including saturatus and sasaimae
Here watkinsorum has replaced watkinsi. It must take this form because Hellmayr intended it to refer to both the Watkins brothers, not just one of them. They were the ones who collected the type specimen. See Costas (2017).
Automolus: The two basal species in Automolus are transferred to Cryptomolus (Claramunt et al. 2013, type rufipileatus).
Buff-throated Foliage-gleaner: In Supplement 59 (2018) the NACC split the Chiriqui Foliage-gleaner, Automolus exsertus, from the Buff-throated Foliage-gleaner, Automolus ochrolaemus. Based on Harvey et al. (2020), the HBW Checklist, and Freeman and Montgomery (2017), I've redone the complex as follows:
- Western Buff-throated Foliage-gleaner, Automolus cervinigularis, including hypophaeus and pallidigularis
- Chiriqui Foliage-gleaner, Automolus exsertus
- Eastern Buff-throated Foliage-gleaner, Automolus ochrolaemus, including turdinus and auricularis
Part of the point is that it seems clear from Freeman and Montgomery that exsertus is vocally different from the Western forms. NACC are clearly of the opinion that it is also different from those to the east, as if on an island (sky island?). Harvey et al. (2020) found 2 million years distance between the Costa Rican birds and those from Loreto, Peru, which are in the ochrolaemus group. This gives us the three species solution above. Yes, it is a bit speculative.
Striped Woodhaunter: Another IOC/Ridgely-Tudor (2009) change is to split Striped Woodhaunter, Automolus subulatus, into Western Woodhaunter, Automolus virgatus and Eastern Woodhaunter, Automolus subulatus.
Notes — Pseudocolaptini
Pacific Tuftedcheek: To improve alignment with the IOC list, I have split the Pacific Tuftedcheek, Pseudocolaptes johnsoni, from the Buffy Tuftedcheek, Pseudocolaptes lawrencii, as advocated by Ridgely and Tudor (2009). The forthcoming article by Forcina, Boesman, and Jowers (2021) shows that all three tuftedcheek species are roughly equally distant from one another.
Notes — Furnariini
Pale-legged Hornero: Another IOC/Ridgely and Tudor (2009) split involves the horneros. Pacific Hornero, Furnarius cinnamomeus, and Caribbean Hornero, Furnarius longirostris, have been split from Pale-legged Hornero, Furnarius leucopus.
Pale-legged Hornero: Based on Harvey et al. (2020) and the HBW Checklist, the Pale-legged Hornero, Furnarius leucopus, is further split into:
- Tricolored Hornero, Furnarius tricolor, including araguaiae and assimilis
- Pale-legged Hornero, Furnarius leucopus
Upucerthia: The genus Upucerthia has been divided into 4 four parts (Chesser et al., 2007; Fjeldså et al. 2007). Two species are moved to Ochetorhynchus (in Pygarrhichadini), which also absorbs Chilia and Eremobius; two others form the new genus Tarphonomus (in Pseudocolaptini; Chesser et al., 2007); U. serrana is placed in the new genus Geocerthia (Chesser et al., 2009); the rest remain in Upucerthia, which shrinks a bit with the lumping of Plain-breasted Earthcreeper, Upucerthia jelskii, into Buff-breasted Earthcreeper, Upucerthia validirostris (Areta and Pearman, 2013). The last two genera remain with the Furnariini.
Cipo Cinclodes: The Cipo Cinclodes, Cinclodes espinhacensis, is treated as a subspecies of the Long-tailed Cinclodes, Cinclodes pabsti. See SACC proposal #548.
Cinclodes: The arrangement of Cinclodes species now follows Harvey et al. (2020), which is similar to Derryberry et al. (2011) and Sanín et al. (2009). Sanín et al. presented evidence for splitting the Bar-winged Cinclodes, Cinclodes fuscus, into three species. Jaramillo (2003) suggested the English names Buff-winged Cinclodes for Cinclodes fuscus and Cream-winged Cinclodes for Cinclodes albiventris. SACC has named Cinclodes albidiventris Chestnut-winged Cinclodes. The Chestnut-winged Cinclodes is the northern group, with a range extending into NW Peru (Cajamarca and Piura). It includes subspecies heterurus and oreobates, in addition to albidiventris. The central species is the Cream-winged Cinclodes, ranging from N Peru (Amazonas) to N Chile (Antofagasta) and NW Argentina (La Rioja). It apparently includes the isolated subspecies riojanus, rufus, and yzurietaeof NW Argentina, as well as tucumanus and albiventris. The southern Buff-winged Cinclodes is then monotypic.
Notes — Synallaxini
Tawny Tit-Spinetail: Following H&M-4 (Dickinson and Christidis, 2014), the Tawny Tit-Spinetail, Sylviorthorhynchus yanacensis, has moved to Sylviorthorhynchus from Leptasthenura. These two sister species are deeply divided from Leptasthenura (Derryberry et al., 2011).
Plain-mantled Tit-Spinetail: Based on Harvey et al. (2020) and the HBW Checklist, the Plain-mantled Tit-Spinetail, Leptasthenura aegithaloides, is split into:
- Plain-mantled Tit-Spinetail, Leptasthenura aegithaloides, including grisescens
- Pallid Tit-Spinetail, Leptasthenura pallida
- Buffy Tit-Spinetail, Leptasthenura berlepschi
Plain Thornbird: The Plain Thornbird, Phacellodomus inornatus has been split from Rufous-fronted Thornbird, Phacellodomus rufifrons, matching IOC and Ridgely and Tudor (2009).
Asthenes: The biggest changes in the Synallaxini involve the genera Asthenes, Oreophylax, and Schizoeaca. Here the usual generic limits do not reflect the gene tree. The recent paper by Derryberry et al. (2010b) has done much to straighten out the situation, and Harvey et al. (2020) did even more.
Four of the Asthenes, humicola, patagonica, steinbachi, and cactorum, do not really belong to the group. Derryberry et al. (2010b) created the new genus Pseudasthenes for this clade. This is a bit surprising as cactorum has been considered a subspecies of the Cordilleran Canastero, Siptornoides modestus, which is definitely not a close relative.
The remaining species of the Asthenes, Oreophylax, and Schizoeaca group form a clade. Derryberry et al. (2010b) suggest treating them all as Asthenes. That's sort of what I do, but using a very narrow conception of Asthenes. But first, we'll look at the broad version of Asthenes, Short-billed Canastero to White-chinned Thistletail.
There are 3 major branches of the broad Asthenes. I separate each as a genus: "Asthenes", Siptornoides, and Asthenes. Generally speaking, the first two groups are canasteros, and the last thistletails. The basal branch includes dorbignyi, baeri, and berlepschi. All of these have previously been considered part of a superspecies. I have separated them and others as "Asthenes" as I have not found an available genus name.
The remaining species of the broad Asthenes are in two clades. The first consists of the birds now in Siptornoides The other, which is comprised of the remaining Asthenes, includes the species formerly placed in Oreophylax and Schizoeaca. These are mostly called thistletails. The number of species involved is rather surprising as all of the old Schizoeaca have been considered conspecific. Note also the geographic separation between the Andean Schizoeaca and Oreophylax (Itatiaia Spinetail) of SE Brazil.
Vilcabamba Thistletail: There is a split in the former Schizoeaca based on Hosner et al. (2015b) and SACC #697. The Ayacucho Thistletail, Asthenes ayacuchensis, has been split from Vilcabamba Thistletail, Asthenes vilcabambae.
Creamy-breasted Canastero: As recommended by IOC and Ridgely and Tudor (2009), Creamy-breasted Canastero, "Asthenes" dorbignyi has been split. Additionally, there is a further split based on Harvey et al. (2020) and the HBW Checklist. This gives us:
- Pale-tailed Canastero, "Asthenes" huancavelicae
- White-tailed Canastero, "Asthenes" usheri
- Rusty-vented Canastero, "Asthenes" dorbignyi
- Dark-winged Canastero, "Asthenes" arequipae
The dorbignyi group include at least one undescribed taxon, the “Ancash” Canastero.
 |
 |
 |
| “Ancash” Canastero | ||
|---|---|---|
Another supposed member of the dorbignyi group, luizae, is on a separate branch that is basal in the Siptornoides clade.
Ash-browed Spinetail: Based on Harvey et al. (2020), the subspecies debilis is sister to the Creamy-crested Spinetail, Cranioleuca albicapilla. As a result, Ash-browed Spinetail, Cranioleuca curtata, is split into:
- Slender-billed Spinetail, Cranioleuca debilis
- Ash-browed Spinetail, Cranioleuca curtata, including cisandina
As you can see from tree, these are not sister taxa. The name Slender-billed has been used in the past for debilis, e.g., by Cory and Hellmayr.
White-bellied Spinetail: The White-bellied Spinetail has been placed in the new genus Mazaria (Claramunt, 2014a) as Mazaria propinqua. SACC still lists it in Synallaxis, but genetic data (Derryberry et al., 2011) showed it was sister to Schoeniophylax and I moved it there. As Claramunt (2014a) notes, the two species are so different that putting the White-bellied Spinetail in Schoeniophylax makes the genus undiagnosible. He established the new genus Mazaria to solve this problem.
Undescribed Spinetail: On a Field Guides trip with Bret Whitney and Marcello Barreiros, Bret pointed out an undescribed Certhiaxis spinetail on a river island. As we were in the Araguaia drainage, this must be the unnamed Certhiaxis mentioned in the Harvey et al. (2020) tree. It's pretty similar to a Yellow-chinned Spinetail, without a yellow chin. I didn't add it to the list, but Harvey et al. (2020) included it in their analysis and you can find it in the species tree next to the Yellow-chinned Spinetail, Certhiaxis cinnamomeus.
Necklaced Spinetail: Based on Harvey et al. (2020), Stopiglia et al. (2020), SACC #882 and the HBW Checklist, the Necklaced Spinetail, Synallaxis stictothorax, is split into:
- Necklaced Spinetail, Synallaxis stictothorax
- Chinchipe Spinetail, Synallaxis chinchipensis
White-lored Spinetail: Harvey et al. (2020) found that a subspecies of Plain-crowned Spinetail, Synallaxis gujanensis is closer to White-lored Spinetail, Synallaxis albilora than to the rest of gujanensis. The race in question is present in Santa Cruz, Boliva, near El Tuná and I infer it is certhiola. Apparently it looks a lot like albilora.
Rufous Spinetail: Based on Harvey et al. (2020) and the HBW Checklist, the Rufous Spinetail, Synallaxis unirufa, is split into:
- Merida Rufous Spinetail, Synallaxis meridana
- Perija Rufous Spinetail, Synallaxis munoztebari
- Colombian Rufous Spinetail, Synallaxis unirufa
- Peruvian Rufous Spinetail, Synallaxis ochrogaster
 |
| Click for Xenopinae thru Furnariini species |
|---|
The Synallaxini have undergone further changes due to Derryberry et al. (2011). There's some reallocation in the Cranioleuca branch. Cranioleuca loses two species, Sulphur-throated Spinetail moves to Limnoctites and Speckled Spinetail moves to Thripophaga. Conversely, Thripophaga loses Russet-mantled Softtail to Cranioleuca. It also gains the newly discovered Delta Amacuro Softtail, Thripophaga amacurensis (Hilty et al., 2013). Cranioleuca also loses a species due to the lumping of Baron's Spinetail, Cranioleuca baroni, into Line-cheeked Spinetail, Cranioleuca antisiensis. See Seeholzer and Brumfield (2018) and SACC Proposal 762 for details.
I had previously separated several species of Synallaxis as Poecilurus in an attempt to avoid paraphyly. The more complete analysis of Derryberry et al. showed that this was a failure. According, I have returned the Poecilurus species to Synallaxis. Moreover, I've also merged the monotypic Gyalophylax and Siptornopsis into Synallaxis. Although Synallaxis is a large genus, the differences between the Synallaxis spinetails are small enough that I see no reason to subdivide it.
Furnariinae: True Ovenbirds G.R. Gray, 1840
Xenopinae: Xenops Bonaparte, 1854
- Streaked Xenops, Xenops rutilans
- Slender-billed Xenops, Xenops tenuirostris
- White-throated Xenops, Xenops minutus
- Northwestern Plain Xenops, Xenops mexicanus
- Southeastern Plain Xenops, Xenops genibarbis
Berlepschiini: Palmcreeper Ohlson et al., 2013a
- Point-tailed Palmcreeper, Berlepschia rikeri
Pygarrhichadini Wolters, 1977
- Rufous-tailed Xenops, Microxenops milleri
- White-throated Treerunner, Pygarrhichas albogularis
- Crag Chilia, Ochetorhynchus melanurus
- Rock Earthcreeper, Ochetorhynchus andaecola
- Straight-billed Earthcreeper, Ochetorhynchus ruficaudus
- Band-tailed Earthcreeper, Ochetorhynchus phoenicurus
Margarornini: Barbtails and Treerunners Ridgway, 1911
- Spotted Barbtail, Premnoplex brunnescens
- White-throated Barbtail, Premnoplex tatei
- Pearled Treerunner, Margarornis squamiger
- Ruddy Treerunner, Margarornis rubiginosus
- Beautiful Treerunner, Margarornis bellulus
- Fulvous-dotted Treerunner / Star-chested Treerunner, Margarornis stellatus
Philydorini: Foliage-gleaners, Treehunters Sclater & Salvin, 1873
- Cryptic Treehunter, Cichlocolaptes mazarbarnetti
- Pale-browed Treehunter, Cichlocolaptes leucophrus
- Cinnamon-rumped Foliage-gleaner, Philydor pyrrhodes
- Sharp-billed Treehunter, Philydor contaminatus
- Black-capped Foliage-gleaner, Philydor atricapillus
- Alagoas Foliage-gleaner, Philydor novaesi
- Slaty-winged Foliage-gleaner, Anabazenops fuscipennis
- Rufous-rumped Foliage-gleaner, Anabazenops erythrocercus
- Dusky-cheeked Foliage-gleaner / Bamboo Foliage-gleaner, Anabazenops dorsalis
- White-collared Foliage-gleaner, Anabazenops fuscus
- Great Xenops, Megaxenops parnaguae
- Ochre-breasted Foliage-gleaner, Anabacerthia lichtensteini
- White-browed Foliage-gleaner, Anabacerthia amaurotis
- Montane Foliage-gleaner, Anabacerthia striaticollis
- Scaly-throated Foliage-gleaner, Anabacerthia variegaticeps
- Rufous-tailed Foliage-gleaner, Anabacerthia ruficaudata
- Buff-browed Foliage-gleaner, Syndactyla rufosuperciliata
- Russet-mantled Foliage-gleaner / Planalto Foliage-gleaner, Syndactyla dimidiata
- White-throated Foliage-gleaner / Tepui Foliage-gleaner, Syndactyla roraimae
- Peruvian Recurvebill, Syndactyla ucayalae
- Bolivian Recurvebill, Syndactyla striata
- Guttulate Foliage-gleaner, Syndactyla guttulata
- Rufous-necked Foliage-gleaner, Syndactyla ruficollis
- Lineated Foliage-gleaner, Syndactyla subalaris
- Chestnut-winged Hookbill, Ancistrops strigilatus
- Buff-fronted Foliage-gleaner, Dendroma rufa
- Chestnut-winged Foliage-gleaner, Dendroma erythroptera
- Chestnut-capped Foliage-gleaner / Henna-capped Foliage-gleaner, Clibanornis rectirostris
- Canebrake Groundcreeper, Clibanornis dendrocolaptoides
- Dusky Foliage-gleaner, Hylocryptus obscurus
- Cinnamon-throated Foliage-gleaner, Hylocryptus cinnamomeigula
- Santa Marta Foliage-gleaner, Hylocryptus rufipectus
- Rusty Foliage-gleaner, Hylocryptus rubiginosus
- Black-tailed Foliage-gleaner, Hylocryptus nigricauda
- Henna-hooded Foliage-gleaner, Hylocryptus erythrocephalus
- Uniform Treehunter, Thripadectes ignobilis
- Flammulated Treehunter, Thripadectes flammulatus
- Rufous-backed Treehunter / Peruvian Treehunter, Thripadectes scrutator
- Striped Treehunter, Thripadectes holostictus
- Streak-breasted Treehunter, Thripadectes rufobrunneus
- Streak-capped Treehunter, Thripadectes virgaticeps
- Black-billed Treehunter, Thripadectes melanorhynchus
- Chestnut-crowned Foliage-gleaner, Cryptomolus rufipileatus
- Brown-rumped Foliage-gleaner, Cryptomolus melanopezus
- Western Buff-throated Foliage-gleaner, Automolus cervinigularis
- Chiriqui Foliage-gleaner, Automolus exsertus
- Eastern Buff-throated Foliage-gleaner, Automolus``` ochrolaemus
- Western Woodhaunter, Automolus virgatus
- Eastern Woodhaunter, Automolus subulatus
- Olive-backed Foliage-gleaner, Automolus infuscatus
- Para Foliage-gleaner, Automolus paraensis
- White-eyed Foliage-gleaner, Automolus leucophthalmus
- Pernambuco Foliage-gleaner, Automolus lammi
Pseudocolaptini Ridgway 1911
- Bolivian Earthcreeper, Tarphonomus harterti
- Chaco Earthcreeper, Tarphonomus certhioides
- Rusty-winged Barbtail, Premnornis guttuliger
- Buffy Tuftedcheek, Pseudocolaptes lawrencii
- Streaked Tuftedcheek, Pseudocolaptes boissonneautii
- Pacific Tuftedcheek, Pseudocolaptes johnsoni
Furnariini G.R. Gray, 1840
- Sharp-tailed Streamcreeper, Lochmias nematura
- Wren-like Rushbird, Phleocryptes melanops
- Curve-billed Reedhaunter, Limnornis curvirostris
- Crested Hornero, Furnarius cristatus
- Lesser Hornero, Furnarius minor
- Rufous Hornero, Furnarius rufus
- Tricolored Hornero, Furnarius tricolor
- Wing-banded Hornero / Band-tailed Hornero, Furnarius figulus
- Caribbean Hornero, Furnarius longirostris
- Pacific Hornero, Furnarius cinnamomeus
- Pale-legged Hornero, Furnarius leucopus
- Pale-billed Hornero / Bay Hornero, Furnarius torridus
- Striated Earthcreeper, Geocerthia serrana
- Buff-breasted Earthcreeper, Upucerthia validirostris
- Patagonian Forest Earthcreeper, Upucerthia saturatior
- White-throated Earthcreeper, Upucerthia albigula
- Scale-throated Earthcreeper / Scaly-throated Earthcreeper, Upucerthia dumetaria
- Stout-billed Cinclodes, Cinclodes excelsior
- Royal Cinclodes, Cinclodes aricomae
- Long-tailed Cinclodes, Cinclodes pabsti
- White-bellied Cinclodes, Cinclodes palliatus
- Dark-bellied Cinclodes, Cinclodes patagonicus
- Surf Cinclodes / Peruvian Seaside Cinclodes, Cinclodes taczanowskii
- Seaside Cinclodes / Chilean Seaside Cinclodes, Cinclodes nigrofumosus
- Buff-winged Cinclodes, Cinclodes fuscus
- Blackish Cinclodes, Cinclodes antarcticus
- Cordoba Cinclodes, Cinclodes comechingonus
- Chestnut-winged Cinclodes, Cinclodes albidiventris
- Cream-winged Cinclodes, Cinclodes albiventris
- White-winged Cinclodes, Cinclodes atacamensis
- Gray-flanked Cinclodes, Cinclodes oustaleti
- Olrog's Cinclodes, Cinclodes olrogi
Synallaxini: Spinetails de Selys-Longchamps, 1839 (1836)
- Masafuera Rayadito, Aphrastura masafuerae
Click for Synallaxini
species (3 pages) - Thorn-tailed Rayadito, Aphrastura spinicauda
- Tawny Tit-Spinetail, Sylviorthorhynchus yanacensis
- Des Murs's Wiretail, Sylviorthorhynchus desmurii
- White-browed Tit-Spinetail, Leptasthenura xenothorax
- Tufted Tit-Spinetail, Leptasthenura platensis
- Araucaria Tit-Spinetail, Leptasthenura setaria
- Striolated Tit-Spinetail, Leptasthenura striolata
- Streaked Tit-Spinetail / Streak-backed Tit-Spinetail, Leptasthenura striata
- Rusty-crowned Tit-Spinetail, Leptasthenura pileata
- Brown-capped Tit-Spinetail, Leptasthenura fuliginiceps
- Plain-mantled Tit-Spinetail, Leptasthenura aegithaloides
- Pallid Tit-Spinetail, Leptasthenura pallida
- Andean Tit-Spinetail, Leptasthenura andicola
- Buffy Tit-Spinetail, Leptasthenura berlepschi
- Lark-like Brushrunner, Coryphistera alaudina
- Firewood-gatherer, Anumbius annumbi
- Streak-fronted Thornbird, Phacellodomus striaticeps
- Little Thornbird, Phacellodomus sibilatrix
- Plain Thornbird, Phacellodomus inornatus
- Rufous-fronted Thornbird, Phacellodomus rufifrons
- Orange-breasted Thornbird, Phacellodomus ferrugineigula
- Orange-eyed Thornbird, Phacellodomus erythrophthalmus
- Greater Thornbird, Phacellodomus ruber
- Freckle-breasted Thornbird, Phacellodomus striaticollis
- Chestnut-backed Thornbird, Phacellodomus dorsalis
- Spot-breasted Thornbird, Phacellodomus maculipectus
- White-browed Spinetail, Hellmayrea gularis
- Short-billed Canastero, "Asthenes" baeri
- Pale-tailed Canastero, "Asthenes" huancavelicae
- White-tailed Canastero, "Asthenes" usheri
- Rusty-vented Canastero, "Asthenes" dorbignyi
- Dark-winged Canastero, "Asthenes" arequipae
- Berlepsch's Canastero, "Asthenes" berlepschi
- Cipo Canastero, Siptornoides luizae
- Hudson's Canastero, Siptornoides hudsoni
- Line-fronted Canastero, Siptornoides urubambensis
- Many-striped Canastero, Siptornoides flammulatus
- Junin Canastero, Siptornoides virgatus
- Scribble-tailed Canastero, Siptornoides maculicauda
- Streak-throated Canastero, Siptornoides humilis
- Cordilleran Canastero, Siptornoides modestus
- Austral Canastero, Siptornoides anthoides
- Streak-backed Canastero, Siptornoides wyatti
- Puna Canastero, Siptornoides sclateri
- Itatiaia Spinetail, Asthenes moreirae
- Sharp-billed Canastero, Asthenes pyrrholeuca
- Maquis Canastero, Asthenes heterura
- Eye-ringed Thistletail, Asthenes palpebralis
- Rusty-fronted Canastero, Asthenes ottonis
- Vilcabamba Thistletail, Asthenes vilcabambae
- Ayacucho Thistletail, Asthenes ayacuchensis
- Puna Thistletail, Asthenes helleri
- Black-throated Thistletail, Asthenes harterti
- Canyon Canastero, Asthenes pudibunda
- Mouse-colored Thistletail, Asthenes griseomurina
- Ochre-browed Thistletail, Asthenes coryi
- Perija Thistletail, Asthenes perijana
- White-chinned Thistletail, Asthenes fuliginosa
- Bay-capped Wren-Spinetail, Spartonoica maluroides
- Rufous Cacholote / Gray-crested Cacholote, Pseudoseisura unirufa
- Caatinga Cacholote, Pseudoseisura cristata
- Brown Cacholote, Pseudoseisura lophotes
- White-throated Cacholote, Pseudoseisura gutturalis
- Pink-legged Graveteiro, Acrobatornis fonsecai
- Orange-fronted Plushcrown, Metopothrix aurantiaca
- Double-banded Graytail, Xenerpestes minlosi
- Equatorial Graytail, Xenerpestes singularis
- Spectacled Prickletail, Siptornis striaticollis
- Speckled Spinetail, Thripophaga gutturata
- Plain Softtail, Thripophaga fusciceps
- Roraiman Barbtail, Thripophaga adusta
- Orinoco Softtail, Thripophaga cherriei
- Delta Amacuro Softtail, Thripophaga amacurensis
- Striated Softtail, Thripophaga macroura
- Straight-billed Reedhaunter, Limnoctites rectirostris
- Sulphur-throated Spinetail / Sulphur-bearded Spinetail, Limnoctites sulphuriferus
- Russet-mantled Softtail, Cranioleuca berlepschi
- Marcapata Spinetail, Cranioleuca marcapatae
- Light-crowned Spinetail, Cranioleuca albiceps
- Rusty-backed Spinetail, Cranioleuca vulpina
- Parker's Spinetail, Cranioleuca vulpecula
- Coiba Spinetail, Cranioleuca dissita
- Scaled Spinetail, Cranioleuca muelleri
- Olive Spinetail, Cranioleuca obsoleta
- Pallid Spinetail, Cranioleuca pallida
- Stripe-crowned Spinetail, Cranioleuca pyrrhophia
- Bolivian Spinetail, Cranioleuca henricae
- Creamy-crested Spinetail, Cranioleuca albicapilla
- Slender-billed Spinetail, Cranioleuca debilis
- Tepui Spinetail, Cranioleuca demissa
- Streak-capped Spinetail, Cranioleuca hellmayri
- Crested Spinetail, Cranioleuca subcristata
- Gray-headed Spinetail, Cranioleuca semicinerea
- Ash-browed Spinetail, Cranioleuca curtata
- Red-faced Spinetail, Cranioleuca erythrops
- Line-cheeked Spinetail, Cranioleuca antisiensis
- Dusky-tailed Canastero, Pseudasthenes humicola
- Patagonian Canastero, Pseudasthenes patagonica
- Cactus Canastero, Pseudasthenes cactorum
- Steinbach's Canastero, Pseudasthenes steinbachi
- White-bellied Spinetail, Mazaria propinqua
- Chotoy Spinetail, Schoeniophylax phryganophilus
- Red-and-white Spinetail, Certhiaxis mustelinus
- Yellow-chinned Spinetail, Certhiaxis cinnamomeus
- Necklaced Spinetail, Synallaxis stictothorax
- Russet-bellied Spinetail, Synallaxis zimmeri
- Chinchipe Spinetail, Synallaxis chinchipensis
- Great Spinetail, Synallaxis hypochondriaca
- Ochre-cheeked Spinetail, Synallaxis scutata
- Gray-bellied Spinetail, Synallaxis cinerascens
- Maranon Spinetail, Synallaxis maranonica
- Plain-crowned Spinetail, Synallaxis gujanensis
- White-lored Spinetail, Synallaxis albilora
- Slaty Spinetail, Synallaxis brachyura
- Silvery-throated Spinetail, Synallaxis subpudica
- Blackish-headed Spinetail, Synallaxis tithys
- Hoary-throated Spinetail, Synallaxis kollari
- Rufous-breasted Spinetail, Synallaxis erythrothorax
- White-whiskered Spinetail, Synallaxis candei
- Rusty-headed Spinetail, Synallaxis fuscorufa
- Black-throated Spinetail, Synallaxis castanea
- Merida Rufous Spinetail, Synallaxis meridana
- Perija Rufous Spinetail, Synallaxis munoztebari
- Colombian Rufous Spinetail, Synallaxis unirufa
- Peruvian Rufous Spinetail, Synallaxis ochrogaster
- Stripe-breasted Spinetail, Synallaxis cinnamomea
- Ruddy Spinetail, Synallaxis rutilans
- Chestnut-throated Spinetail, Synallaxis cherriei
- Dusky Spinetail, Synallaxis moesta
- Cabanis's Spinetail, Synallaxis cabanisi
- McConnell's Spinetail, Synallaxis macconnelli
- Pinto's Spinetail, Synallaxis infuscata
- Rufous-capped Spinetail, Synallaxis ruficapilla
- Bahia Spinetail, Synallaxis cinerea
- Red-shouldered Spinetail, Synallaxis hellmayri
- Cinereous-breasted Spinetail, Synallaxis hypospodia
- Spix's Spinetail, Synallaxis spixi
- Dark-breasted Spinetail, Synallaxis albigularis
- Rio Orinoco Spinetail, Synallaxis beverlyae
- Pale-breasted Spinetail, Synallaxis albescens
- Azara's Spinetail, Synallaxis azarae
- Apurimac Spinetail, Synallaxis courseni
- Sooty-fronted Spinetail, Synallaxis frontalis